Research Area: Biochemistry, Biophysics, and Structural Biology
Structural intuitions lead to structural insights
Jennifer Viegas | PNAS
November 8, 2024
HHMI Investigator and Professor of Biology and Chemistry Catherine Drennan has spent a distinguished career addressing challenging and wide-ranging structural biology problems.
Catherine Drennan, a Howard Hughes Medical Institute investigator and professor and a professor of biology and chemistry at the Massachusetts Institute of Technology (MIT), has spent a distinguished career addressing challenging and wide-ranging structural biology problems. These include her discovery, while she was a graduate student, of the structure of vitamin B12 bound to protein and her recent determination at atomic resolution of the structure of an active ribonucleotide reductase (RNR) with water molecules, findings reported in her Inaugural Article (IA) (1).
Drennan, who was elected to the National Academy of Sciences in 2023, has uncovered the form and function of metalloenzymes that use metal cofactors to catalyze chemical reactions involving free radicals. Metalloenzymes are of broad human health and environmental interest; some are promising antibiotic and cancer drug targets, whereas others hold the potential for bioremediation efforts, such as converting carbon dioxide into biofuels.
Family of Accomplished Scientists
Drennan was raised in New York City by her father, an obstetrician–gynecologist, and her mother, an anthropologist. Her father was born in Germany and attended medical school at the University of Hamburg. Harboring antifascist leanings, he fled Germany in 1933. He completed medical training in Geneva, Switzerland, before obtaining political asylum in 1940 in the United States, where he became one of the first doctors to practice the Lamaze Method of natural childbirth.
Drennan’s mother attended Antioch College, where she was a student of civil engineer Arthur Ernest Morgan, who was appointed in 1948 to India’s first University Education Commission. She accompanied Morgan to India and served as his administrative assistant before earning her doctorate in anthropology at Cornell University.
“Both my parents were endlessly curious,” says Drennan. “My father was fascinated by the molecular basis of medicine, and my mother was fascinated by people and instilled in me her love for storytelling, teaching, and mentoring.”
Diagnosed with Dyslexia
Although she was an attentive student, Drennan did not learn to read until her second time through sixth grade. “When I finally learned how to read, it was by memorizing the shapes of words,” says Drennan, who was diagnosed with dyslexia when she was in first grade. “Over time I became very good at shape recognition. I am not disabled; I am differently abled. The skill set that I developed to compensate for my dyslexia has made me a world-class structural biologist. We all have strengths and weaknesses, and my ‘weakness’ is also my superpower.”
She was accepted to Vassar College, where she earned a bachelor’s degree in chemistry in 1985. “Miriam Rossi was my undergraduate chemistry research advisor, and she believed in me before I believed in me,” Drennan says. Upon Rossi’s advice, Drennan pursued a doctorate, but not before teaching high-school science and drama at Scattergood Friends School in Iowa.
Following three years of high-school teaching, Drennan pursued graduate studies at the University of Michigan, Ann Arbor, where she earned a PhD in 1995, served as a research fellow from 1995 to 1996, and was mentored by biochemists Martha Ludwig and Rowena Matthews. “They treated me as a colleague, allowing me to see myself as a scientist of value,” Drennan says. “I learned so much from these two incredible scientists. They are, and always will be, my heroes.”
Structure of Vitamin B12 Bound to Protein
With Ludwig et al., Drennan determined the structure of cobalamin (vitamin B12) bound to protein (2). This crystal structure revealed how the protein modulates the reactivity of the B12 cofactor to enable its critical roles in metabolism.
From 1996 to 1999, Drennan did a postdoctoral stint at the California Institute of Technology, under the mentorship of structural biologist Douglas Rees. “Doug taught by example that one does not have to be cutthroat to succeed in the competitive area of structural biology,” she says. “He has continued to mentor me throughout my career, helping me through challenging times.”
Another important mentor was chemist JoAnne Stubbe, a leader in the study of RNRs who recruited Drennan to MIT in 1999 as an assistant professor of chemistry and has been her collaborator for the past 25 years. Drennan says, “Her passion for scientific discovery is unmatched and has inspired me to keep digging to try to understand, at the most fundamental level, how ribonucleotide reductase works.” Drennan advanced to an associate professorship at MIT in 2004 and a full professorship in 2006.
Revealing Metalloenzyme Form and Function
Drennan’s group continues to study B12 and has provided numerous snapshots of cobalamin-dependent proteins and protein complexes. The findings have changed what is known about B12 functions and mechanisms. Using X-ray crystallography, the researchers unveiled a protein complex capable of methyl transfer from folate to B12 (3). They obtained snapshots of the biological process involved in loading B12 into an enzyme (4) and provided structural data on how B12 can be repurposed from enzyme cofactor to light sensor (5).
Drennan has also worked on uncovering the structures of enzymes containing radical S-adenosylmethionine (SAM) cofactors. Drennan and colleagues revealed an X-ray structure of a radical SAM enzyme (6), helping to establish the “core” fold for an enzyme superfamily that has over 100,000 members. Her group further elucidated structures of SAM family members with functions including posttranslational modification (7), antibiotic and antiviral compound biosynthesis (8, 9), and vitamin biosynthesis (6, 10).
Mononuclear nonheme iron enzymes are also of interest to Drennan. The cofactor is simple, but the reactions catalyzed are complex. Her group reported the structure of a nonheme iron halogenase, showing that the halogen binds directly to the catalytic iron (11). Drennan says, “This was a complete surprise that required new mechanistic proposals to be written.”
“Oceanic Methane Paradox”
Early in her independent career, Drennan determined one of the first structures of a nickel-iron-sulfur-dependent carbon monoxide dehydrogenase (CODH), which plays an important role in the global carbon cycle (12). The structure, along with that of an associated enzyme complex (13), provided a series of snapshots of the multiple metal ion centers underlying the ability of certain microbes to live off hydrogen gas and carbon dioxide in a process known as acetogenesis. More recently, they investigated the molecular basis by which the activity of CODH enzymes can be restored after oxygen exposure (14), a discovery with implications for the industrial use of CODHs.
Drennan and her team have also studied the organic compound methylphosphonate that was proposed as a source of methane from the aerobic upper ocean; the biological source was long a mystery. When a methylphosphonate synthase was discovered by chemical biologist Wilfred van der Donk and coworkers, part of the mystery was solved but the gene for this enzyme did not appear to be widespread. When Drennan and colleagues solved the structure of a methylphosphonate synthase; however, they discovered a sequence motif showing that the gene was, in fact, abundant in microbes that inhabit the upper ocean (15). This seminal finding is credited with resolving the oceanic methane paradox.
Radical-Based Chemistry in Ribonucleotide Reductases
Human RNR is an established chemotherapeutic target, and bacterial RNRs hold promise as antibiotic targets. So Drennan and her team have a longstanding interest in uncovering the mechanisms of RNRs. In 2002, her lab determined the structure of a B12-dependent RNR, which showed how cobalamin could be used to initiate radical chemistry (16). Nearly a decade later, Drennan’s team revealed how high levels of the nucleotide deoxyadenosine triphosphate (dATP) down-regulate RNR activity (17, 18). They subsequently provided structures showing the molecular basis of allosteric specificity, which maintains the proper ratios of RNA to DNA building blocks (19), and demonstrated the importance of RNR activity regulation (20).
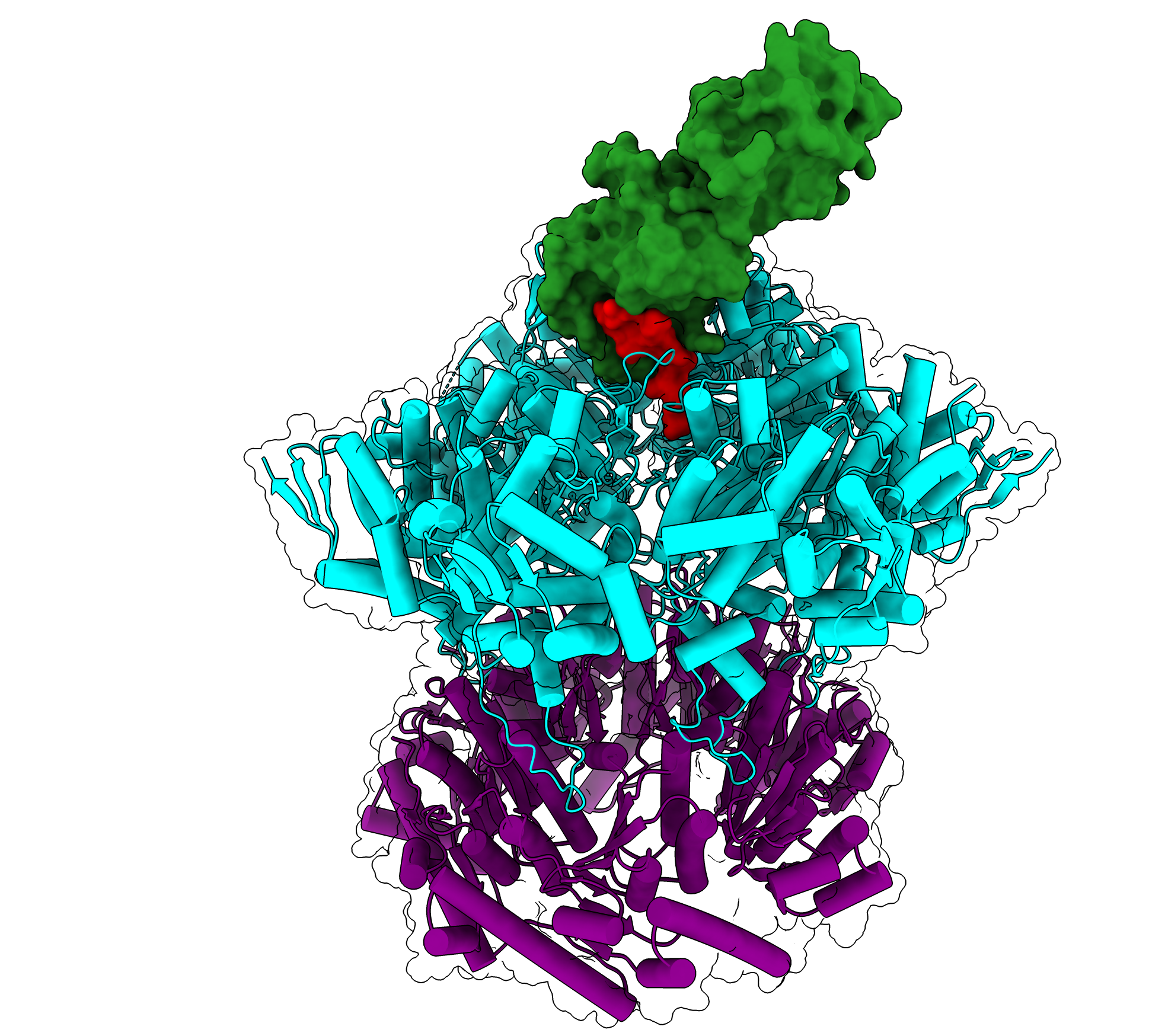
An atomic-resolution structure of any RNR in an active state had been elusive for many years. Drennan and her team achieved the feat in 2020 when they trapped the active state of Escherichia coli RNR and determined its structure by cryoelectron microscopy (21). However, the resolution of the structure was too low for the visualization of water molecules believed to be critical in the radical transfer pathway.
In her IA, Drennan (1) describes how her team resolved the problem, presenting the structure of an active RNR at atomic resolution allowing for the visualization of water molecules. She explains, “This time, instead of using unnatural amino acids to trap the structure, we used a mechanism-based inhibitor. It was a very long road to get to these data, but it was worth the wait.”
“Superheroes of the Cell”
For her achievements, Drennan has received MIT’s Everett Moore Baker Memorial Award for Excellence in Undergraduate Teaching (2005, 2024), the Dorothy Crowfoot Hodgkin Award from The Protein Society (2020), and the William C. Rose Award from the American Society for Biochemistry and Molecular Biology (2023), among other honors. She has mentored nearly 100 undergraduates and dozens of graduate students and postdoctoral associates, many of whom are from underrepresented minority groups or disadvantaged backgrounds. She considers her students extended family members and takes pride in their accomplishments.
She and her team continue to work on RNR using the tools of structural biology. She says, “We want to obtain a deeper level of understanding of the human RNR, which is a cancer drug target. We also want to identify differences between the human enzyme and bacterial RNRs, differences that could be exploited in the development of new antibiotics.”
Beyond these efforts, Drennan’s overall goal is to understand how enzymes control radical species to enable challenging chemical reactions without damaging themselves or their cellular environment. “Radical enzymes are like the Avengers, powerful but with a high potential for collateral damage,” she explains. “I am fascinated by how nature catalyzes the most challenging of chemical reactions. The enzymes that do this work are the superheroes of the cell and I want to know their secrets.”
1.
D. E. Westmoreland et al., 2.6-Å resolution cryo-EM structure of a class Ia ribonucleotide reductase trapped with mechanism-based inhibitor N3CDP. Proc. Natl. Acad. Sci. U.S.A.121, e2417157121 (2024). Crossref. PubMed.
2.
C. L. Drennan et al., How a protein binds B12: A 3.0 Å X-ray structure of B12-binding domains of methionine synthase. Science266, 1669–1674 (1994). Crossref. PubMed.
3.
Y. Kung et al., Visualizing molecular juggling within a B12-dependent methyltransferase complex. Nature484, 265–269 (2012). Crossref. PubMed.
4.
F. A. Vaccaro, D. A. Born, C. L. Drennan, Structure of metallochaperone in complex with the cobalamin-binding domain of its target mutase provides insight into cofactor delivery. Proc. Natl. Acad. Sci. U.S.A.120, e2214085120 (2023). Crossref. PubMed.
5.
M. Jost et al., Structural basis for gene regulation by a B12-dependent photoreceptor. Nature526, 536–541 (2015). Crossref. PubMed.
6.
F. Berkovitch et al., Crystal structure of biotin synthase, an S-Adenosylmethionine-dependent radical enzyme. Science303, 76–79 (2004). Crossref. PubMed.
7.
J. L. Vey et al., Structural basis for glycyl radical formation by pyruvate formate-lyase activating enzyme. Proc. Natl. Acad. Sci. U.S.A.105, 16137–16141 (2008). Crossref. PubMed.
8.
J. Bridwell-Rabb et al., A B12-dependent radical SAM enzyme involved in oxetanocin A biosynthesis. Nature544, 322–326 (2017). Crossref. PubMed.
9.
P. J. Goldman, T. L. Grove, S. J. Booker, C. L. Drennan, X-ray analysis of butirosin biosynthetic enzyme BtrN redefines structural motifs for AdoMet radical chemistry. Proc. Natl. Acad. Sci. U.S.A.40, 15949–15954 (2013). Crossref.
10.
M. I. McLaughlin et al., Crystallographic snapshots of sulfur insertion by lipoyl synthase. Proc. Natl. Acad. Sci. U.S.A.113, 9446–9450 (2016). Crossref. PubMed.
11.
L. C. Blasiak, F. H. Vaillancourt, C. T. Walsh, C. L. Drennan, Crystal structure of the non-haem iron halogenase SyrB2 in syringomycin biosynthesis. Nature440, 368–371 (2006). Crossref. PubMed.
12.
C. L. Drennan et al., Life on carbon monoxide: X-ray structure of Rhodospirillum rubrum Ni-Fe-S carbon monoxide dehydrogenase. Proc. Natl. Acad. Sci. U.S.A.98, 11973–11978 (2001). Crossref. PubMed.
13.
T. I. Doukov et al., A Ni-Fe-Cu center in a bifunctional carbon monoxide dehydrogenase/acetyl-CoA synthase. Science298, 567–572 (2002). Crossref. PubMed.
14.
E. C. Wittenborn et al., Redox-dependent rearrangements of the NiFeS cluster of carbon monoxide dehydrogenase. Elife7, e39451 (2018). Crossref. PubMed.
15.
D. A. Born et al., Structural basis for methylphosphonate biosynthesis. Science358, 1336–1339 (2017). Crossref. PubMed.
16.
M. D. Sintchak et al., The crystal structure of class II ribonucleotide reductase reveals how an allosterically regulated monomer mimics a dimer. Nat. Struct. Mol. Biol.9, 293–300 (2002). Crossref.
17.
N. Ando et al., Structural interconversions modulate activity of Escherichia coli ribonucleotide reductase. Proc. Natl. Acad. Sci. U.S.A.108, 21046–21051 (2011). Crossref. PubMed.
18.
N. Ando et al., Allosteric inhibition of human ribonucleotide reductase by dATP entails the stabilization of a hexamer. Biochemistry55, 373–381 (2016). Crossref. PubMed.
19.
C. M. Zimanyi et al., Molecular basis for allosteric specificity regulation in class Ia ribonucleotide reductase from Escherichia coli. Elife5, e07141 (2016). Crossref. PubMed.
20.
P.Y.-T. Chen, M. A. Funk, E. J. Brignole, C. L. Drennan, Disruption of an oligomeric interface prevents allosteric inhibition of Escherichia coli class Ia ribonucleotide reductase. J. Biol. Chem.293, 10404–10412 (2018). Crossref. PubMed.
21.
G. Kang, A. T. Taguchi, J. Stubbe, C. L. Drennan, Structure of a trapped radical transfer pathway within a ribonucleotide reductase holocomplex. Science368, 424–427 (2020). Crossref. PubMed.
Rest in pieces: deconstructing polypeptide degradation machinery
Lillian Eden | Department of Biology
November 12, 2024
Research from the Sauer and Davis Labs in the Department of Biology at MIT shows that conformational changes contribute to the specificity of “molecular woodchippers”
Degradation is a crucial process for maintaining protein homeostasis by culling excess or damaged proteins whose components can then be recycled. It is also a highly regulated process—for good reason. A cell could potentially waste many resources if the degradation machinery destroys proteins it shouldn’t.
One of the major pathways for protein degradation in bacteria and eukaryotic mitochondria involves a molecular machine called ClpXP. ClpXP is made up of two components: a star-shaped structure made up of six subunits called ClpX that engages and unfolds proteins tagged for degradation, and an associated barrel-shaped enzyme, called ClpP, that chemically breaks up proteins into small pieces called peptides.
ClpXP is incredibly adaptable and is often compared to a woodchipper — able to take in materials and spit out their broken-down components. Thanks to biochemical experiments, this molecular degradation machine is known to be able to break down hundreds of different proteins in the cell regardless of physical or chemical properties such as size, shape, or charge. ClpX uses energy from ATP hydrolysis to unfold proteins before they are threaded through its central channel, referred to as the axial channel, and into the degradation chamber of ClpP.
In three papers, one in PNAS and two in NatureCommunications, researchers from the Department of Biology at MIT have expanded our understanding of how this molecular machinery engages with, unfolds, and degrades proteins — and how that machinery refrains, by design, from unfolding proteins not tagged for degradation.
Alireza Ghanbarpour, until recently a postdoc in the Sauer Lab and Davis Lab and first author on all three papers, began with a simple question: given the vast repertoire of potential substrates — that is, proteins to be degraded — how is ClpXP so specific?
Ghanbarpour approached the question of ClpXP’s versatility by characterizing conformational changes of the molecular machine using a technique called cryogenic electron microscopy. In cryo-EM, sample particles are frozen in solution, and images are collected; algorithms then create 3D renderings from the 2D images.
“It’s really useful to generate different structures in different conditions and then put them together until you know how a machine works,” he says. “I love structural biology, and these molecular machines make fascinating targets for structural work and biochemistry. Their structural plasticity and precise functions offer exciting opportunities to understand how nature leverages enzyme conformations to generate novel functions and tightly regulate protein degradation within the cell.”
Inside the cell, these proteases do not work alone but instead work together with “adaptor” proteins, which can promote — or inhibit — degradation by ClpXP. One of the adaptor proteins that promotes degradation by ClpXP is SspB.
In E. coli and most other bacteria, ClpXP and SspB interact with a tag called ssrA that is added to incomplete proteins when their biosynthesis on ribosomes stalls.
The tagging process frees up the ribosome to make more proteins, but creates a problem: incomplete proteins are prone to aggregation, which could be detrimental to cellular health and can lead to disease. By interacting with the degradation tag, ClpXP and SspB help to ensure the degradation of these incomplete proteins. Understanding this process and how it may go awry may open therapeutic avenues in the future.
“It wasn’t clear how certain adapters were interacting with the substrate and the molecular machines during substrate delivery,” Ghanbarpour notes. “My recent structure reveals that the adapter engages with the enzyme, reaching deep into the axial channel to deliver the substrate.”
Ghanbarpour and colleagues showed that ClpX engages with both the SspB adaptor and the ssrA degradation tag of an ill-fated protein at the same time. Surprisingly, they also found that this interaction occurs while the upper part of the axial channel through ClpX is closed — in fact, the closed channel allows ClpX to contact both the tag and the adaptor simultaneously.
This result was surprising, according to senior author andSalvador E. Luria Professor of Biology Robert Sauer, whose lab has been working on understanding this molecular machine for more than two decades: it was unclear whether the channel through ClpX closes in response to a substrate interaction, or if the channel is always closed until it opens to pass an unfolded protein down to ClpP to be degraded.
Preventing Rogue Degradation
Throughout this project, Ghanbarpour was co-advised by structural biologist and Associate Professor of Biology Joey Davis and collaborated with members of the Davis Lab to better understand the conformational changes that allow these molecular machines to function. Using a cryo-EM analysis approach developed in the Davis lab called CryoDRGN, the researchers showed that there is an equilibrium between ClpXP in the open and closed states: it’s usually closed but is open in about 10% of the particles in their samples.
The closed state is almost identical to the conformation ClpXP assumes when it is engaged with an ssrA-tagged substrate and the SspB adaptor.
To better understand the biological significance of this equilibrium, Ghanbarpour created a mutant of ClpXP that is always in the open position. Compared to normal ClpXP, the mutant degraded some proteins lacking obvious degradation tags faster but degraded ssrA-tagged proteins more slowly.
According to Ghanbarpour, these results indicate that the closed channel improves ClpXP’s ability to efficiently engage tagged proteins meant to be degraded, whereas the open channel allows more “promiscuous” degradation.
Pausing the Process
The next question Ghanbarpour wanted to answer was what this molecular machine looks like while engaged with a protein it is attempting to unfold. To do that, he created a substrate with a highly stable protein attached to the degradation tag that is initially pulled into ClpX, but then dramatically slows protein unfolding and degradation.
In the structures where the degradation process stalls, Ghanbarpour found that the degradation tag was pulled far into the molecular machine—through ClpX and into ClpP—and the folded protein part of the substrate was pulled tightly against the axial channel of ClpX.
The opening of the axial channel, called the axial pore, is made up of looping protein structures called RKH loops. These flexible loops were found to play roles both in recognizing the ssrA degradation tag and in how substrates or the SspB adaptor interact with or are pulled against the channel during degradation.
The flexibility of these RKH loops allows ClpX to interact with a large number of different proteins and adapters, and these results clarify some previous biochemical and mutational studies of interactions between the substrate and ClpXP.
Although Ghanbarpour’s recent work focused on just one adaptor and degradation tag, he noted there are many more targets — ClpXP is something akin to a Swiss army knife for breaking down polypeptide chains.
The way those other substrates interact with ClpXP could differ from the structures solved with the SspB adaptor and ssrA tag. It also stands to reason that the way ClpXP reacts to each substrate may be unique. For example, given that ClpX is occasionally in an open state, some substrates may engage with ClpXP only while it’s in an open conformation.
In his new position at Washington University, Ghanbarpour intends to continue exploring how ClpXP and other molecular machines locate their target substrates and interact with adaptors, shedding light on how cells regulate protein degradation and maintain protein homeostasis.
The structures Ghanbarpour solved involved free-floating protein degradation machinery, but membrane-bound degradation machinery also exists. The membrane-bound version’s structure and conformational adaptions potentially differ from the structures Ghanbarpour found in his previous three papers. Indeed, in a recent preprint, Ghanbarpour worked on the cryo-EM structure of a nautilus shell-shaped protein assembly that seems to control membrane-bound degradation machinery. This assembly plays a critical role in regulating protein degradation within the bacterial inner membrane.
“The function of these proteases goes beyond simply degrading damaged proteins. They also target transcription factors, regulatory proteins, and proteins that don’t exist in normal conditions,” he says. “My new lab is particularly interested in understanding how cells use these proteases and their accessory adaptors, both under normal and stress conditions, to reshape the proteome and support recovery from cellular distress.”
Cells make variants of thousands of proteins. These variants are not produced indiscriminately, but rather through precise regulatory mechanisms that can meet rapidly changing needs of the cell according to new research from the Cheeseman Lab.
Greta Friar | Whitehead Institute
October 18, 2024
Our cells contain thousands of proteins that have gone largely undetected and unstudied until recent years: these are variants of known proteins, which cells can make when their protein-building machinery interacts differently with the same stretch of genetic code. These protein variants have typically been overlooked as occasional accidents of gene expression, but researchers including Whitehead Institute Member Iain Cheeseman are discovering that they are actually abundant and can play important roles in cell functions. Researchers in Cheeseman’s lab are studying individual protein variants to learn more about them and their roles in health and disease, but they also wanted to understand broader patterns of protein variant production: how do cells control when to make one variant of a protein versus another, and what are the consequences of such switches?
Cheeseman, who is also a professor of biology at the Massachusetts Institute of Technology, and graduate student in his lab Jimmy Ly have now identified how cells switch to a different pattern of protein variant production during mitosis, or cell division. In research published in the journal Nature on October 23, they show that this broad regulatory switch helps cells survive paused cell divisions that can sometimes occur in healthy humans or be triggered by certain chemotherapy treatments. The work confirms that cells make variants of thousands of proteins, and also demonstrates that cells do not do so indiscriminately. Rather, cells use precise regulatory mechanisms to switch between different patterns of protein variant production, in order to rapidly tailor the proteins available to fit the changing needs of the cell.
A plethora of hidden proteins
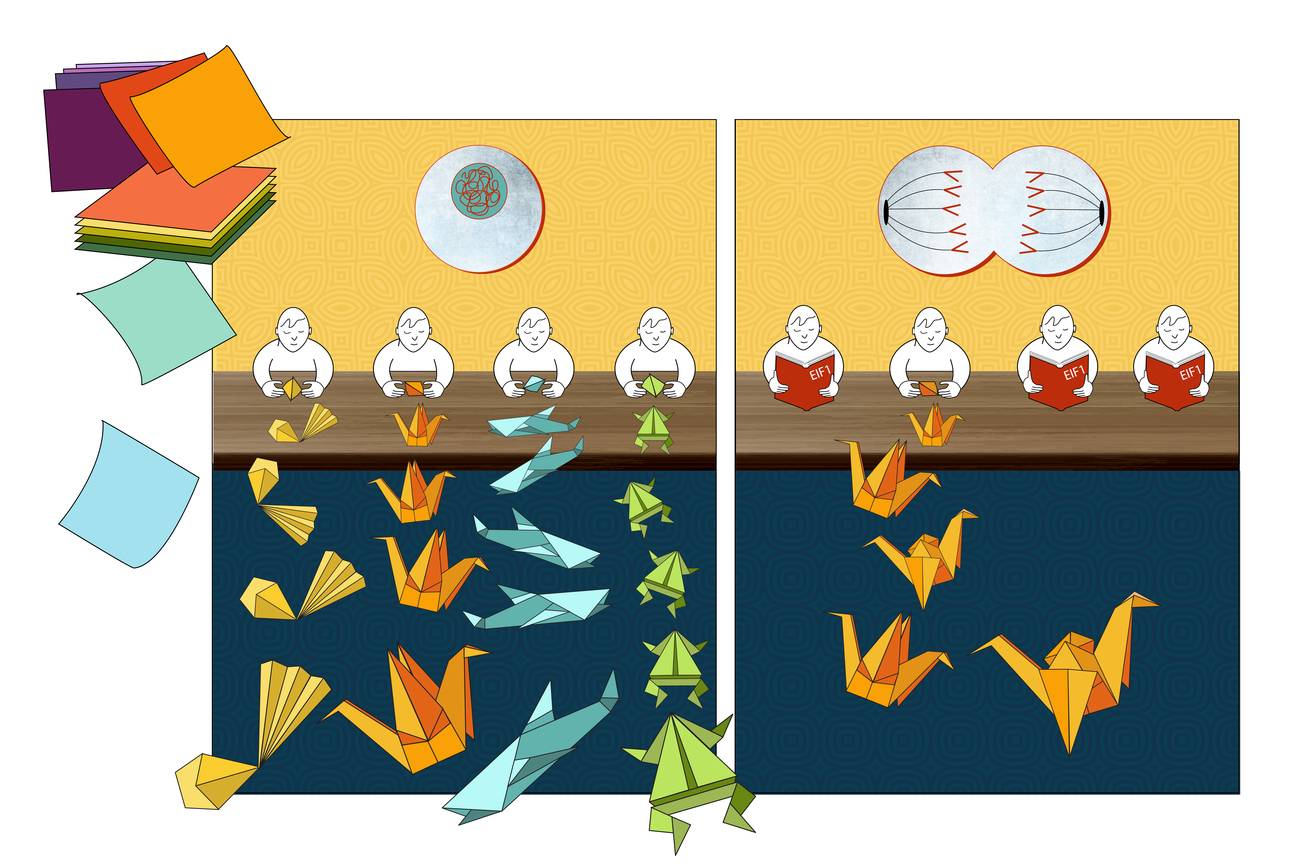
Hw can our cells contain unknown proteins? In high school biology classes, students learn the rule that each gene codes for exactly one protein, such that if you know an organism’s genetic code, you should know every protein it can make. In fact, there are instead many genes that code for multiple proteins. For a protein to be made, first the genetic code for it is copied from DNA into a messenger RNA (mRNA). Then, a ribosome, the cellular machine that follows the instructions in genetic code to build a protein, locates the coding sequence within the mRNA by scanning for the start codon, a sequence of the three bases A, U, and G – bases are the chemical building blocks of RNA, abbreviated as A, U, C, and G. The ribosome recognizes the AUG start codon as the place to begin following instructions, and builds a protein based on the genetic sequence from there through to another trio of bases called a stop codon. However, one way that different versions of a protein can be produced is that a ribosome may begin reading the instructions from multiple different starting points.
Sometimes, a ribosome may miss the first AUG start codon and skip ahead to another AUG somewhere in the middle of the gene’s code, creating a truncated version of the protein. Sometimes, a ribosome may treat a similar trio of bases, such as CUG or GUG, as a start codon. This can cause it to begin earlier, creating a protein based on an extended genetic sequence. These possibilities mean that cells contain thousands more different proteins, or variants of proteins, than are represented by the dogma of one gene, one protein.
In order to understand protein variant production, the researchers—in collaboration with researchers from Whitehead Institute Member David Bartel’s lab–used a method that let them carefully track ribosomes to compare which start sites ribosomes tended to use. They looked at start site selection during mitosis versus during the rest of the cell cycle and found that a dramatic shift in use occurred for thousands of start sites. Specifically, the researchers found that during mitosis, ribosome scanning becomes more stringent. The ribosome will only begin making proteins at AUG sequences, and even then, only at AUGs that have preferable sequences of bases surrounding them—known as a strong Kozak context. This increased selectivity does not always lead to the familiar version of the protein being made during mitosis; sometimes the first AUG start codon has a weak Kozak context, so a truncated protein gets made from an AUG start codon with a stronger Kozak context that lies within the gene.
“Coming into this project, we knew very little about protein production during mitosis—for a long time, people didn’t think much protein production happened in mitosis at all,” Ly says. “It was satisfying to show not only that it is occurring, but that there’s a shift in which proteins are being made—and that this shift is important for cellular viability.”
How cells switch between protein variant programs
The researchers next identified how the switch to increased stringency is initiated during mitosis. They discovered that the key player is a protein called eIF1, which is one of many partners that can pair with ribosomes to help them select their start site. In particular, increased eIF1 pairing with ribosomes causes the ribosomes to be more stringent in their start codon selection, inhibiting the usage of non-AUG initiation sites or sites with weak Kozak contexts.
During mitosis, ribosome pairing with eIF1 increases sharply, leading to the shift in stringency. This change in pairing rate during mitosis puzzled the researchers: ribosomes and their partners, including eIF1, all typically reside together in the main body of the cell—where ribosomes make proteins—so they should be able to pair freely at any time. The researchers looked for other molecules in the same location that could be altering how ribosomes and eIF1 interact during different parts of the cell cycle, but they couldn’t find anything. Eventually, the researchers realized that the answer to the puzzle lay in a separate location: the nucleus.
They found that cells maintain a large pool of eIF1 inside of the nucleus, locked away from the ribosomes. Then, during cell division, the wall of the nucleus dissolves, mixing its contents with the rest of the cell. This is necessary for the dividing cell to divvy up its DNA, but it also releases the pool of eIF1 to pair with ribosomes, increasing stringency. At the end of mitosis, the nucleus reforms and eIF1 is re-incorporated into the nucleus of each of the two daughter cells, and the cells return to a less stringent program.
“The explanation for increased interaction between eIF1 and ribosomes during mitosis had really stumped us, and so when I saw eIF1 localizing to the nucleus, that was a really exciting ‘aha’ moment,” Ly says. “Discovering this mechanism of nuclear release during mitosis was unexpected, and it’s interesting to think about how else cells might be using it.”
Consequences of increased stringency for the cell
Once the researchers understood the how, they then wanted to understand the why? What they discovered is that when cells have no nuclear pool of eIF1, and so no change in stringency during mitosis, they are more likely to die during mitosis. In particular, these cells fare poorly during mitotic arrest, a state in which cells get stuck in mitosis for hours or even days–much longer than typical mitosis. Arrest occurs when cells detect a possible cell division error and so halt their division until the error is corrected or the cell dies.
One effect of increased stringency during mitosis is related to mitochondria, which are required for energy production in many cell types and are therefore required for maintaining viability. Cells stuck in mitotic arrest need energy to keep them going through this unexpected delay. The researchers found that increased stringency during mitosis led to an increase in the production of important mitochondrial proteins, boosting the cells’ energy supply to get them through arrest.
Increased stringency also gives cells the tools they need to escape arrest, even if they haven’t fixed the error that caused them to pause division. In a Nature paper in 2023, Cheeseman and then-postdoc in his lab Mary-Jane Tsang showed that when cells build up enough of the truncated version of a protein called CDC20, they can escape arrest. Ly’s work adds to this story by showing that the nuclear release of eIF1 increases stringency, leading to more production of truncated CDC20 during mitosis, which explains how cells build up enough of this protein variant during mitosis to trigger their escape. These findings may have important potential implications for some cancer chemotherapy strategies.
Some chemotherapies work by trapping cancer cells in mitotic arrest until they die. Cheeseman, Tsang, and Ly’s work collectively shows that when cancer cells lack sufficient truncated CDC20—as can occur in the absence of nuclear eIF1—the cells cannot escape arrest and so are killed off by these chemotherapies at higher rates. These results could be used to improve the efficacy of antimitotic chemotherapy drugs.
The switch in protein variant production that the researchers found affects thousands of proteins. These newly identified protein variants serve as a foundation for many future projects in the lab.
As the researchers continue to examine the consequences of this switch to stringency during mitosis, they are also searching for other cases in which cells regulate protein variant production outside of mitosis. For example, the researchers are interested in how this switch in stringency affects fertility; immature egg cells spend a long time in a form of arrested cell division without an intact nucleus, and Ly observed eIF1 in the nucleus of the immature female eggs.
“Cells have axes of control that they use to quickly make broad changes in gene expression,” Cheeseman says. “Several of these are central to controlling cell division—for example, the role of phosphorylation as a regulatory switch in mitosis has been well studied. Our work identifies another axis of control, and we’re excited to discover more about when and how cells make use of it.”
Killing the messenger
Lillian Eden | Department of Biology
October 23, 2024
Newly characterized anti-viral defense system in bacteria aborts infection through novel mechanism by chemically modifying mRNA.
Like humans and other complex multicellular organisms, single-celled bacteria can fall ill and fight off viral infections. A bacterial virus is known as a bacteriophage, or, more simply, a phage, which is one of the most ubiquitous life forms on Earth. Phages and bacteria are engaged in a constant battle, the virus attempting to circumvent the bacteria’s defenses, and the bacteria racing to find new ways to protect itself.
These anti-phage defense systems are carefully controlled and prudently managed — dormant but always poised to strike.
New research recently published in Naturefrom the Laub Lab in the Department of Biology at MIT has characterized an anti-phage defense system in bacteria known as CmdTAC. CmdTAC prevents viral infection by altering mRNA, the single-stranded genetic code used to produce proteins, of both the host and the virus.
This defense system detects phage infection at a stage when the viral phage has already commandeered the host’s machinery for its own purposes. In the face of annihilation, the ill-fated bacterium activates a defense system that will halt translation, preventing the creation of new proteins and aborting the infection — but dooming itself in the process.
“When bacteria are in a group, they’re kind of like a multicellular organism that is not connected to one another. It’s an evolutionarily beneficial strategy for one cell to kill itself to save another identical cell,” says Christopher Vassallo, a postdoc and co-author of the study. “You could say it’s like self-sacrifice: one cell dies to protect the other cells.”
The enzyme responsible for altering the mRNA is called an ADP-ribosyltransferase. Researchers have characterized hundreds of these enzymes — although only a few are known to target DNA or other types of RNA, all but a handful target proteins. This is the first time these enzymes have been characterized targeting mRNA within cells.
Expanding understanding of anti-phage defense
Co-first author and graduate student Chris Doering noted that it is only within the last decade or so that researchers have begun to appreciate the breadth of diversity and complexity of anti-phage defense systems. For example, CRISPR gene editing, a technique used in everything from medicine to agriculture, is rooted in research on the bacterial CRISPR-Cas9 anti-phage defense system.
CmdTAC is a subset of a widespread anti-phage defense mechanism called a toxin-antitoxin system. A TA system is just that: a toxin capable of killing or altering the cell’s processes rendered inert by an associated antitoxin.
Although these TA systems can be identified — if the toxin is expressed by itself, it kills or inhibits the growth of the cell; if the toxin and antitoxin are expressed together, the toxin is neutralized — characterizing the cascade of circumstances that activates these systems requires extensive effort. In recent years, however, many TA systems have been shown to serve as anti-phage defenses.
Two general questions need to be answered to understand a viral defense system: how do bacteria detect an infection, and how do they respond?
Detecting infection
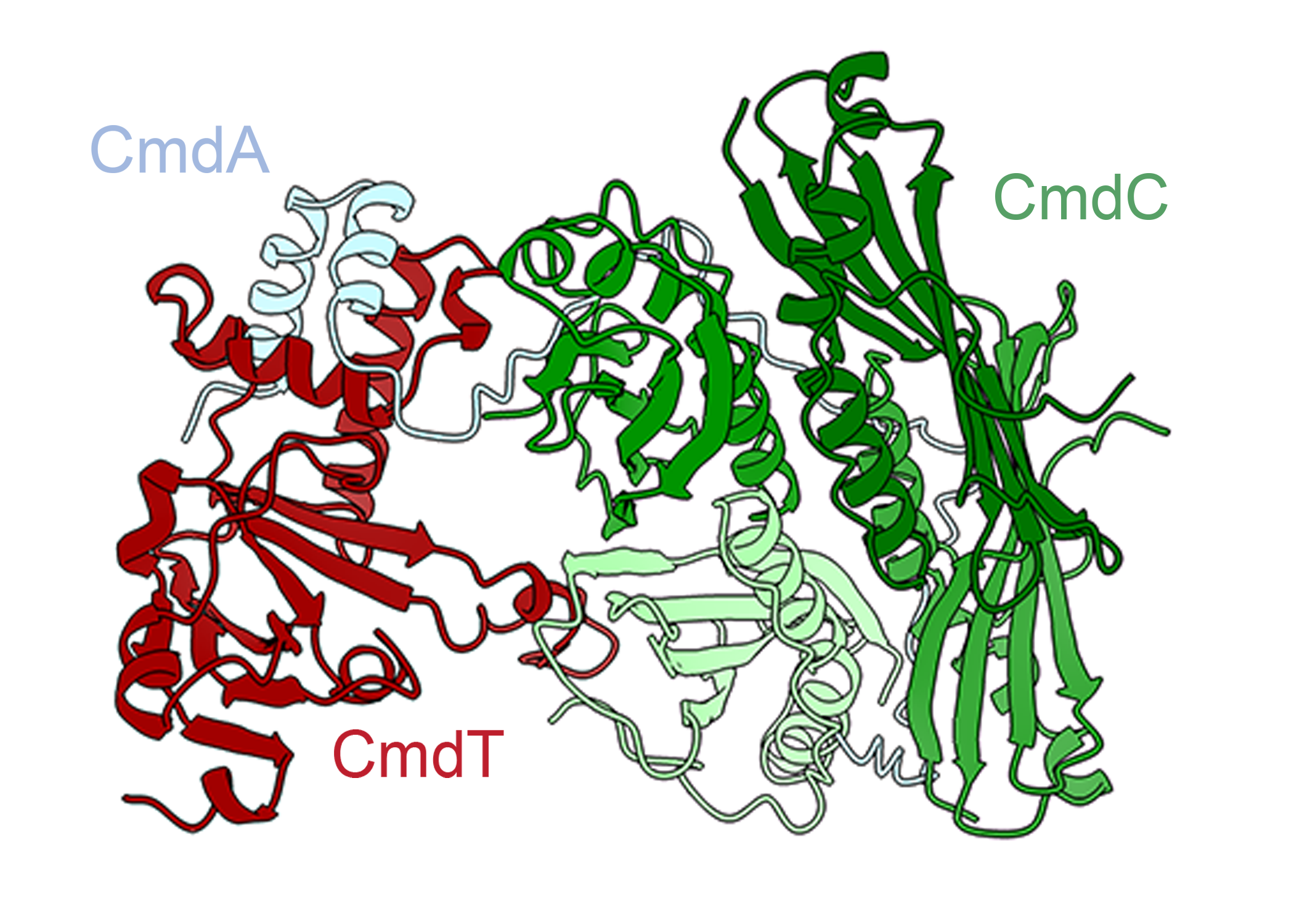
CmdTAC is a TA system with an additional element, and the three components generally exist in a stable complex: the toxin CmdT, the antitoxin CmdA, and an additional component that mediates the system, the chaperone CmdC.
If the phage’s protective capsid protein is present, CmdC disassociates from CmdT and CmdA and interacts with the phage capsid protein instead. In the model outlined in the paper, the chaperone CmdC is, therefore, the sensor of the system, responsible for recognizing when an infection is occurring. Structural proteins, such as the capsid that protects the phage genome, are a common trigger because they’re abundant and essential to the phage.
The uncoupling of CmdC leads to the degradation of the neutralizing antitoxin CmdA, which releases the toxin CmdT to do its lethal work.
Toxicity on the loose
Guided by computational tools, the researchers knew that CmdT was likely an ADP-ribosyltransferase due to its similarities to other such enzymes. As the name suggests, the enzyme transfers an ADP ribose onto its target.
To determine how CmdT was altering mRNA, the researchers tested a mix of short sequences of single-stranded RNA to see if the enzyme was drawn to any sequences or positions in particular. RNA has four bases: A, U, G, and C, and the evidence points to the enzyme recognizing GA sequences.
The CmdT modification of GA sequences in mRNA blocks its translation. The cessation of creating new proteins aborts the infection, preventing the phage from spreading beyond the host to infect other bacteria.
“Not only is it a new type of bacterial immune system, but the enzyme involved does something that’s never been seen before: the ADP-ribsolyation of mRNA,” Vassallo says.
Although the paper outlines the broad strokes of the anti-phage defense system, there’s more to learn: it’s unclear how CmdC interacts with the capsid protein, and how the chemical modification of GA sequences prevents translation.
Beyond Bacteria
While exploring anti-phage defense aligns with the Laub Lab’s overall goal of understanding how bacteria function and evolve, these results may have broader implications beyond bacteria.
Senior author Michael Laub, Salvador E. Luria Professor and HHMI Investigator, says the ADP-ribosyltransferase has homologs in eukaryotes, including human cells. They are not well studied, and not currently among the Laub Lab’s research topics, but they are known to be up-regulated in response to viral infection.
“There are so many different — and cool — mechanisms by which organisms defend themselves against viral infection,” Laub says. “The notion that there may be some commonality between how bacteria defend themselves and how humans defend themselves is a tantalizing possibility.”
New snapshots of ancient life
Alison Biester | Department of Chemistry
October 3, 2024
The resolution revolution, beating “blobology”, and shedding light on how ancient microbes thrived in a primordial soup.
The earliest life on earth created biological molecules despite the limited materials available in the primordial soup such as CO2, hydrogen gas, and minerals containing iron, nickel, and sulfur.
As ancient microbes evolved, they developed proteins that sped up chemical reactions, called enzymes. Enzymes were evolutionarily advantageous because they created local environments called active sites optimized for reaction performance.
Although we know that carbon is the building block of life on earth–we wouldn’t exist without carbon-based molecules such as proteins and DNA–much remains unclear about how more complex carbon-based molecules were originally generated from CO2. Proteins and DNA are huge molecules with thousands of carbon atoms, so creating life from CO2 would be no small undertaking.
Catherine Drennan, Professor of Biology and Chemistry and HHMI Investigator and Professor, has long studied the enzymes that perform these crucial reactions wherein CO2 is converted into a form of carbon that cells can use, which requires iron, nickel, and sulfur.
In particular, she uses structural biology to study carbon monoxide dehydrogenase (CODH), which reacts with CO2 to produce CO, and acetyl-CoA synthase (ACS), which uses CO with another single unit of carbon to create a carbon-carbon bond. Crystallographic work by Drennan and others has provided structural snapshots of bacterial CODH and ACS, but its structure in other contexts remains elusive. During my PhD, I worked with Drennan on the structural characterization of CODH and ACS, culminating in a publication in PNAS, published October 3, 2024.
Throughout Drennan’s career, the lab has used a method known as X-ray crystallography to determine enzyme structures at atomic resolution. In recent years, however, cryogenic electron microscopy (cryo-EM) has risen in popularity as a structural biology technique.
Cryo-EM offers some key advantages over X-ray crystallography, such as its ability to capture structures of large and dynamic complexes. However, cryo-EM is limited in its ability to elucidate structures of small proteins, an area where X-ray crystallography continues to excel.
To perform a cryo-EM experiment, proteins are rapidly frozen in a thin layer of ice and imaged on an electron microscope. By capturing images of the protein in various orientations, researchers can generate a 3D model of their protein of interest.
Around 2015, cryo-EM reached a tipping point known as the “resolution revolution.” Due to improvements in both the hardware for collecting cryo-EM data and the software used for data processing, the technique could, for the first time, be used to determine protein structures at near-atomic resolution.
Seeing the potential for this new technique, MIT opened its very own cryo-EM facility with two electron microscopes in 2018. Just a year later, I joined the Drennan lab. When I began my thesis work, Cathy asked “Would you like to do crystallography or cryo-EM?”
Eager to try something that was both novel for researchers and new to me, I chose cryo-EM.
Ancient microbes
An ancient type of microbe, archaea, also uses CODH and ACS. Without information on how these protein chains interact, we cannot understand how these proteins work together within this complex–but it’s a difficult question to answer. In total, the complex contains forty protein chains that interact with one another and adopt various conformations to perform their chemistry.
We don’t know for sure which ACS enzyme came first, the bacterial or archaeal one, but we know they are both very ancient.
Archaeal CODH has been visualized via X-ray crystallography, but that CODH was isolated from the enormous megadalton enzyme complex present in the native archaea.
A CO2 molecule, which reacts with CODH, is 44 daltons; the enzyme complex at 2.2 megadaltons is 50,000 times the size of CO2. The complex consists of several copies of CODH, ACS, and a cobalt-containing enzyme that donates the second one-carbon unit used by ACS. Due to the large and dynamic nature of the complex, it was a great candidate for visualizing with cryo-EM.
Just before his retirement, Grahame grew hundreds of liters of archaea and isolated approximately one gram of the enzyme complex that he provided to the Drennan Lab for structural characterization. Each cryo-EM experiment can use as little as a microgram of protein. For a structural biologist, having one gram of protein–in theory, enough for one million experiments–to work with is a dream.
Blobology
With an abundance of protein, I embarked on this project with this exciting new technique on a promising target. I prepared my cryo-EM sample and collected data at the new MIT cryo-EM facility. As I was collecting data, I could see in the images large protein complexes that appeared to be my complex of interest. I could also see some smaller proteins that were consistent with the shape of isolated CODH. When I went on to process my data, I focused on the larger protein complexes, since the structure of isolated CODH was already known.
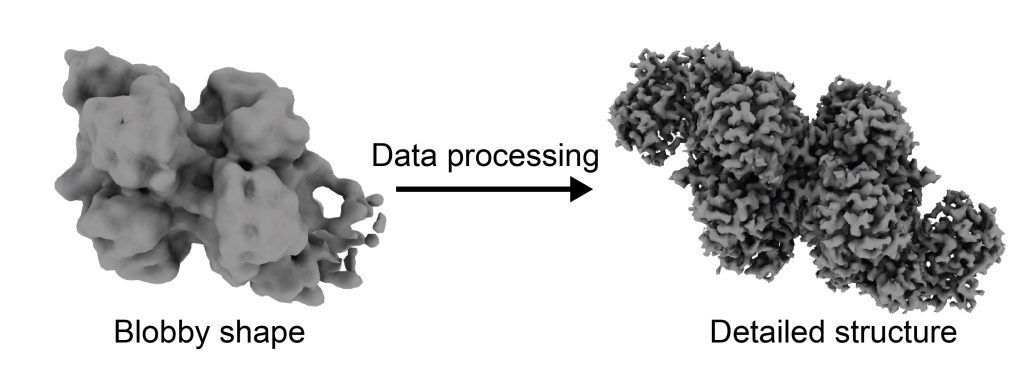
However, when I finished processing my first dataset, I was a bit disappointed. My resolution was very low–instead of atoms, I was seeing amorphous blobs, and I had no idea which blob matched with which protein, or how the proteins fit together. Rather than post-revolution cryo-EM, I felt like I was performing the “blobology” of the past.
Our cryo-EM data contains detailed structural information that becomes evident after significant data processing. On the left is the initial structure of our proteins of interest, carbon monoxide dehydrogenase (CODH) and acetyl-CoA synthase (ACS), and on the right is our final, detailed one. Photo courtesy of Alison Biester.
But the project was young, and a few failed experiments are par for the course of a PhD.
The next step was sample optimization, and luckily I had plenty of sample to work with. I tried preparing the protein in a different way, changed the protein concentration, used different additives, and scaled up my data collection.
Nothing helped. No matter what I tried, I could not move out of blobology territory. So, as one does when a project is failing, I stepped away. I worked on other projects and stopped thinking about the archaeal CODH and ACS.
A few months later, the cryo-EM facility was seeking users to try a new sample preparation instrument called the chameleon. Chameleon automates the sample preparation process and is intended to improve sample quality. With plenty of sample still to spare, I volunteered to try the instrument.
Just prior to my data collection, the facility had also installed a new software that allows data processing as it is being collected. The software uses automated processes to select proteins within your data; previously, I had only selected large protein complexes consistent with my complex of interest after the fact.
The new software is not very discriminating–but I was surprised when I looked at the results of the live processing. The processing showed that I had a protein complex in my sample that I did not expect – a complex of CODH and ACS!
This complex had just one copy of CODH and one copy of ACS, unlike the full complex that has multiple copies of each. My excitement for the project was reinvigorated. With this new target, could I leave blobology behind and finally join the resolution revolution?
After running more experiments and collecting more data and a few months of data processing, I realized that the sample contained three different states: isolated CODH, CODH with one copy of ACS, and CODH with two copies of ACS. I was able to use the Model-based Analysis of Volume Ensembles (MAVEn) tool developed by the Davis Lab at MIT to sort out these three states. When I finished the data processing, I achieved near-atomic resolution of all three states.
Through this work, for the first time, we can see what the archaeal ACS looks like. The archaeal ACS is fundamentally different from the bacterial one: a huge portion of the enzyme is missing, including part of the enzyme that makes up the active site in bacteria, leaving open the question of what the ACS active site looks like in archaea.
In our structure of archaeal ACS in complex with CODH, we were surprised to see that the active site looks almost identical to the bacterial one. This similarity is enabled by the archaeal CODH, which compensates for the missing part of ACS.
Given how similar the ACS active site environment in bacterial and archaea, we are likely getting a look at an active site that has remained conserved over billions of years of evolution.
Although the project didn’t fulfill its original promise of solving the structure of the large, dynamic protein complex, I did find intriguing insights. The tools available in 2015 would not have enabled me to achieve these results; it is clear to me that the resolution revolution is far from over, and the evolution of structural biology has been fascinating to experience. Cryo-EM has and will continue to evolve, as amazing new tools are still being developed.
Since graduating from MIT, I’ve been working at the Protein Data Bank, the data center that houses all available protein structure information. Working here gives me a front-row view of new discoveries in structural biology. I’m so excited to see where this field will go in the future.
Study reveals the drug, 5-fluorouracil, acts differently in different types of cancer — a finding that could help researchers design better drug combinations.
Anne Trafton | MIT News
October 7, 2024
Since the 1950s, a chemotherapy drug known as 5-fluorouracil has been used to treat many types of cancer, including blood cancers and cancers of the digestive tract.
Doctors have long believed that this drug works by damaging the building blocks of DNA. However, a new study from MIT has found that in cancers of the colon and other gastrointestinal cancers, it actually kills cells by interfering with RNA synthesis.
The findings could have a significant effect on how doctors treat many cancer patients. Usually, 5-fluorouracil is given in combination with chemotherapy drugs that damage DNA, but the new study found that for colon cancer, this combination does not achieve the synergistic effects that were hoped for. Instead, combining 5-FU with drugs that affect RNA synthesis could make it more effective in patients with GI cancers, the researchers say.
“Our work is the most definitive study to date showing that RNA incorporation of the drug, leading to an RNA damage response, is responsible for how the drug works in GI cancers,” says Michael Yaffe, a David H. Koch Professor of Science at MIT, the director of the MIT Center for Precision Cancer Medicine, and a member of MIT’s Koch Institute for Integrative Cancer Research. “Textbooks implicate the DNA effects of the drug as the mechanism in all cancer types, but our data shows that RNA damage is what’s really important for the types of tumors, like GI cancers, where the drug is used clinically.”
Yaffe, the senior author of the new study, hopes to plan clinical trials of 5-fluorouracil with drugs that would enhance its RNA-damaging effects and kill cancer cells more effectively.
Jung-Kuei Chen, a Koch Institute research scientist, and Karl Merrick, a former MIT postdoc, are the lead authors of the paper, which appears today in Cell Reports Medicine.
An unexpected mechanism
Clinicians use 5-fluorouracil (5-FU) as a first-line drug for colon, rectal, and pancreatic cancers. It’s usually given in combination with oxaliplatin or irinotecan, which damage DNA in cancer cells. The combination was thought to be effective because 5-FU can disrupt the synthesis of DNA nucleotides. Without those building blocks, cells with damaged DNA wouldn’t be able to efficiently repair the damage and would undergo cell death.
Yaffe’s lab, which studies cell signaling pathways, wanted to further explore the underlying mechanisms of how these drug combinations preferentially kill cancer cells.
The researchers began by testing 5-FU in combination with oxaliplatin or irinotecan in colon cancer cells grown in the lab. To their surprise, they found that not only were the drugs not synergistic, in many cases they were less effective at killing cancer cells than what one would expect by simply adding together the effects of 5-FU or the DNA-damaging drug given alone.
“One would have expected that these combinations to cause synergistic cancer cell death because you are targeting two different aspects of a shared process: breaking DNA, and making nucleotides,” Yaffe says. “Karl looked at a dozen colon cancer cell lines, and not only were the drugs not synergistic, in most cases they were antagonistic. One drug seemed to be undoing what the other drug was doing.”
Yaffe’s lab then teamed up with Adam Palmer, an assistant professor of pharmacology at the University of North Carolina School of Medicine, who specializes in analyzing data from clinical trials. Palmer’s research group examined data from colon cancer patients who had been on one or more of these drugs and showed that the drugs did not show synergistic effects on survival in most patients.
“This confirmed that when you give these combinations to people, it’s not generally true that the drugs are actually working together in a beneficial way within an individual patient,” Yaffe says. “Instead, it appears that one drug in the combination works well for some patients while another drug in the combination works well in other patients. We just cannot yet predict which drug by itself is best for which patient, so everyone gets the combination.”
These results led the researchers to wonder just how 5-FU was working, if not by disrupting DNA repair. Studies in yeast and mammalian cells had shown that the drug also gets incorporated into RNA nucleotides, but there has been dispute over how much this RNA damage contributes to the drug’s toxic effects on cancer cells.
Inside cells, 5-FU is broken down into two different metabolites. One of these gets incorporated into DNA nucleotides, and other into RNA nucleotides. In studies of colon cancer cells, the researchers found that the metabolite that interferes with RNA was much more effective at killing colon cancer cells than the one that disrupts DNA.
That RNA damage appears to primarily affect ribosomal RNA, a molecule that forms part of the ribosome — a cell organelle responsible for assembling new proteins. If cells can’t form new ribosomes, they can’t produce enough proteins to function. Additionally, the lack of undamaged ribosomal RNA causes cells to destroy a large set of proteins that normally bind up the RNA to make new functional ribosomes.
The researchers are now exploring how this ribosomal RNA damage leads cells to under programmed cell death, or apoptosis. They hypothesize that sensing of the damaged RNAs within cell structures called lysosomes somehow triggers an apoptotic signal.
“My lab is very interested in trying to understand the signaling events during disruption of ribosome biogenesis, particularly in GI cancers and even some ovarian cancers, that cause the cells to die. Somehow, they must be monitoring the quality control of new ribosome synthesis, which somehow is connected to the death pathway machinery,” Yaffe says.
New combinations
The findings suggest that drugs that stimulate ribosome production could work together with 5-FU to make a highly synergistic combination. In their study, the researchers showed that a molecule that inhibits KDM2A, a suppressor of ribosome production, helped to boost the rate of cell death in colon cancer cells treated with 5-FU.
The findings also suggest a possible explanation for why combining 5-FU with a DNA-damaging drug often makes both drugs less effective. Some DNA damaging drugs send a signal to the cell to stop making new ribosomes, which would negate 5-FU’s effect on RNA. A better approach may be to give each drug a few days apart, which would give patients the potential benefits of each drug, without having them cancel each other out.
“Importantly, our data doesn’t say that these combination therapies are wrong. We know they’re effective clinically. It just says that if you adjust how you give these drugs, you could potentially make those therapies even better, with relatively minor changes in the timing of when the drugs are given,” Yaffe says.
He is now hoping to work with collaborators at other institutions to run a phase 2 or 3 clinical trial in which patients receive the drugs on an altered schedule.
“A trial is clearly needed to look for efficacy, but it should be straightforward to initiate because these are already clinically accepted drugs that form the standard of care for GI cancers. All we’re doing is changing the timing with which we give them,” he says.
The researchers also hope that their work could lead to the identification of biomarkers that predict which patients’ tumors will be more susceptible to drug combinations that include 5-FU. One such biomarker could be RNA polymerase I, which is active when cells are producing a lot of ribosomal RNA.
The research was funded by the Damon Runyon Cancer Research Fund, a Ludwig Center at MIT Fellowship, the National Institutes of Health, the Ovarian Cancer Research Fund, the Holloway Foundation, and the STARR Cancer Consortium.
The enemy within: new research reveals insights into the arsenal Rickettsia parkeri uses against its host
Lillian Eden | Department of Biology
July 29, 2024
Identifying secreted proteins is critical to understanding how obligately intracellular pathogens hijack host machinery during infection, but identifying them is akin to finding a needle in a haystack.
For then-graduate student Allen Sanderlin, PhD ’24, the first indication that a risky, unlikely project might work was cyan, tic tac-shaped structures seen through a microscope — proof that his bacterial pathogen of interest was labeling its own proteins.
Sanderlin, a member of the Lamason Lab in the Department of Biology at MIT, studies Rickettsia parkeri, a less virulent relative of the bacterial pathogen that causes Rocky Mountain Spotted Fever, a sometimes severe tickborne illness. No vaccine exists and definitive tests to diagnose an infection by Rickettsia are limited.
Rickettsia species are tricky to work with because they are obligately intracellular pathogens whose entire life cycles occur exclusively inside cells. Many approaches that have advanced our understanding of other bacterial infections and how those pathogens interact with their host aren’t applicable to Rickettsia because they can’t be grown on a plate in a lab setting.
In a paper recently published in Nature Communications, the Lamason Lab outlines an approach for labeling and isolating R. parkeri proteins released during infection. This research reveals seven previously unknown secreted factors, known as effectors, more than doubling the number of known effectors in R. parkeri.
Better-studied bacteria are known to hijack the host’s machinery via dozens or hundreds of secreted effectors, whose roles include manipulating the host cell to make it more susceptible to infection. However, finding those effectors in the soup of all other materials within the host cell is akin to looking for a needle in a haystack, with an added twist that researchers aren’t even sure what those needles look like for Rickettsia.
Approaches that worked to identify the six previously known secreted effectors are limited in their scope. For example, some were found by comparing pathogenic Rickettsia to nonpathogenic strains of the bacteria, or by searching for proteins with domains that overlap with effectors from better-studied bacteria. Predictive modeling, however, relies on proteins being evolutionarily conserved.
“Time and time again, we keep finding that Rickettsia are just weird — or, at least, weird compared to our understanding of other bacteria,” says Sanderlin, the paper’s first author. “This labeling tool allows us to answer some really exciting questions about rickettsial biology that weren’t possible before.”
The cyan tic tacs
To selectively label R. parkeri proteins, Sanderlin used a method called cell-selective bioorthogonal non-canonical amino acid tagging. BONCAT was first described in research from the Tirrell Lab at Caltech. The Lamason Lab, however, is the first group to use the tool successfully in an obligate intracellular bacterial pathogen; the thrilling moment when Sanderlin saw cyan tic-tac shapes indicated successfully labeling only the pathogen, not the host.
Sanderlin next used an approach called selective lysis, carefully breaking open the host cell while leaving the pathogen, filled with labeled proteins, intact. This allowed him to extract proteins that R. parkeri had released into its host because the only labeled proteins amid other host cell material were effectors the pathogen had secreted.
Sanderlin had successfully isolated and identified seven needles in the haystack, effectors never before identified in Rickettsia biology. The novel secreted rickettsial factors are dubbed SrfA, SrfB, SrfC, SrfD, SrfE, SrfF, and SrfG.
“Every grad student wants to be able to name something,” Sanderlin says. “The most exciting — but frustrating — thing was that these proteins don’t look like anything we’ve seen before.”
Special delivery
Theoretically, Sanderlin says, once the effectors are secreted, they work independently from the bacteria — a driver delivering a pizza does not need to check back in with the store at every merge or turn.
Since SrfA-G didn’t resemble other known effectors or host proteins the pathogen could be mimicking during infection, Sanderlin then tried to answer some basic questions about their behavior. Where the effectors localize, meaning where in the cell they go, could hint at their purpose and what further experiments could be used to investigate it.
To determine where the effectors were going, Sanderlin added the effectors he’d found to uninfected cells by introducing DNA that caused human cell lines to express those proteins. The experiment succeeded: he discovered that different Srfs went to different places throughout the host cells.
SrfF and SrfG are found throughout the cytoplasm, whereas SrfB localizes to the mitochondria. That was especially intriguing because its structure is not predicted to interact with or find its way to the mitochondria, and the organelle appears unchanged despite the presence of the effector.
Further, SrfC and SrfD found their way to the endoplasmic reticulum. The ER would be especially useful for a pathogen to appropriate, given that it is a dynamic organelle present throughout the cell and has many essential roles, including synthesizing proteins and metabolizing lipids.
Aside from where effectors localize, knowing what they may interact with is critical. Sanderlin showed that SrfD interacts with Sec61, a protein complex that delivers proteins across the ER membrane. In keeping with the theme of the novelty of Sanderlin’s findings, SrfD does not resemble any proteins known to interact with the ER or Sec61.
With this tool, Sanderlin identified novel proteins whose binding partners and role during infection can now be studied further.
“These results are exciting but tantalizing,” Sanderlin says. “What Rickettsia secrete — the effectors, what they are, and what they do is, by and large, still a black box.”
There are very likely other effectors in the proverbial cellular haystack. Sanderlin found that SrfA-G are not found in every species of Rickettsia, and his experiments were solely conducted with Rickettsia at late stages of infection — earlier windows of time may make use of different effectors. This research was also carried out in human cell lines, so there may be an entirely separate repertoire of effectors in ticks, which are responsible for spreading the pathogen.
Expanding Tool Development
Becky Lamason, the senior author of the Nature Communications paper, noted that this tool is one of a few avenues the lab is exploring to investigate R. parkeri, including a paper in the Journal of Bacteriology on conditional genetic manipulation. Characterizing how the pathogen behaves with or without a particular effector is leaps and bounds ahead of where the field was just a few years ago when Sanderlin was Lamason’s first graduate student to join the lab.
“What I always hoped for in the lab is to push the technology, but also get to the biology. These are two of what will hopefully be a suite of ways to attack this problem of understanding how these bacteria rewire and manipulate the host cell,” Lamason says. “We’re excited, but we’ve only scratched the surface.”
Small changes in the molecular machines that carry out RNA interference can lead to big differences in the efficacy of gene silencing. These new findings from the Bartel Lab have implications for the design of gene-silencing therapeutics.
Greta Friar | Whitehead Institute
July 17, 2024
RNA interference (RNAi) is a process that many organisms, including humans, use to decrease the activity of target RNAs in cells by triggering their degradation or slicing them in half. If the target is a messenger RNA, the intermediary between gene and protein, then RNAi can decrease or completely silence expression of the gene. Researchers figured out how to tailor RNAi to target different RNAs, and since then it has been used as a research tool to silence genes of interest. RNAi is also used in a growing number of therapeutics to silence genes that contribute to disease.
However, researchers still do not understand some of the biochemistry underlying RNAi. Slight differences in the design of the RNAi machinery can lead to big differences in how effective it is at decreasing gene expression. Through trial and error, researchers have worked out guidelines for making the most effective RNAi tools without understanding exactly why they work. However, Whitehead Institute Member David Bartel and graduate student in his lab Peter Wang have now dug deeper to figure out the mechanics of the main cellular machine involved in RNAi. The researchers’ findings, shared in Molecular Cell on July 17, not only provide explanations for some of the known rules for RNAi tool design, but also provide new insights that could improve future designs.
Slicing speed is highly variable
The cellular machine that carries out RNAi has two main parts. One is a guide RNA, a tiny RNA typically only 22 bases or nucleotides long. RNA, like DNA, is made of four possible bases, although RNA has the base uracil (U) instead of the DNA base thymine (T). RNA bases bind to each other in certain pairings—guanines (G) pair to cytosines (C) and adenines (A) pair to U’s—and the sequence of bases in the guide RNA corresponds to a complementary sequence within the target RNA. When the guide RNA comes across a target, the corresponding bases pair up, binding the RNAs. Then the other part of the RNAi machine, an Argonaute protein bound to the guide RNA, can slice the target RNA in half or trigger the cell to break it down more gradually.
In humans, AGO2 is the Argonaute protein that is best at slicing. Only a couple dozen RNA targets actually get sliced, but these few targets play essential roles in processes such as neuron signal control and accurate body shape formation. Slicing is also important for RNAi tools and therapeutics.
In order for AGO2 to slice its target, the target must be in the exact right position. As the guide and target RNAs bind together, they go through a series of motions to ultimately form a double helix. Only in that configuration can AGO2 slice the target.
Researchers had assumed that AGO2 slices through different target RNAs at roughly the same rate, because most research into this process used the same few guide RNAs. These guide RNAs happen to have similar features, and so similar slicing kinetics—but they turn out not to be representative of most guide RNAs.
Wang paired AGO2 with a larger variety of guide RNAs and measured the rate at which each AGO2-guide RNA complex sliced its targets. He found big differences. Whereas the commonly used guide RNAs might differ in their slicing rate by 2-fold, the larger pool of guide RNAs differed by as much as 250-fold. The slicing rates were often much slower than the researchers expected. Previously, researchers thought that all targets could be sliced relatively quickly, so the rate wasn’t considered as a limiting factor – other parts of the process were thought to determine the overall pace – but Wang found that slicing can sometimes be the slowest step.
“The important consideration is whether the slicing rate is faster or slower than other processes in the cell,” Wang says. “We found that for many guide RNAs, the slicing rate was the limiting factor. As such, it impacted the efficacy.”
The slower AGO2 is to slice targets, the more messenger RNAs will remain intact to be made into protein, meaning that the corresponding gene will continue being expressed. The researchers observed this in action: the guide RNAs with slower slicing rates decreased target gene expression by less than the faster ones.
Small changes lead to big differences in slicing rate
Next, the researchers explored what could be causing such big differences in slicing rate between guide RNAs. They mutated guide RNAs to swap out single bases along the guide RNA’s sequence—say, switching the 10th base in the sequence from a C to an A—and measured how this changed the slicing rate.
“The important consideration is whether the slicing rate is faster or slower than other processes in the cell,” Wang says. “We found that for many guide RNAs, the slicing rate was the limiting factor. As such, it impacted the efficacy.”
The researchers found that slicing rate increased when the base at position 7 was an A or a U. The bases A and U pair more weakly than C and G. The researchers found that having a weak A-U pair at that position, or a fully mismatched pair at position 6 or 7, may allow a kink to form in the double helix shape that actually makes the target easier to slice. Wang also found that slicing rate increases with certain substitutions at the 10th and the 17th base positions, although the researchers could not yet determine possible underlying mechanisms.
These observations correspond to existing recommendations for RNAi design, such as not using a G at position 7. The new work demonstrates that the reason these recommendations work is because they affect the slicing rate, and, in the case of position 7, the new work further identifies the specific mechanism at play.
Interplay between regions matters
People designing synthetic guide RNAs thought that the bases at the tail end, past the 16th position, were not very important. This is because in the case of the most commonly used guide RNAs, the target will be rapidly cleaved even if all of the tail end positions are mismatches that cannot pair.
However, Wang and Bartel found that the identity of the tail end bases are only irrelevant in a specific scenario that happens to be true of the most commonly used guide RNAs: when the bases in the center of the guide RNA (positions 9-12) are strong-pairing Cs and Gs. When the center pairings are weak, then the tail end bases need to be perfect matches to the target RNA. The researchers found that guide RNAs could have up to a 600-fold difference in tolerance for tail end mismatches based on the strength of their central pairings.
The reason for this difference has to do with the final set of motions that the two RNAs must perform in order to assume their final double helix shape. A perfectly paired tail end makes it easier for the RNAs to complete these motions. However, a strong enough center can pull the RNAs into the double helix even if the tail ends are not ideally suited for doing so.
The observation that weak central pairing requires perfect or near perfect tail end matches could provide a useful new guideline for designing synthetic RNAs. Any guide RNA runs the risk of sometimes binding other messenger RNAs that are similar enough to the intended target RNA. In the case of a therapy, this off-target binding can lead to negative side effects. Bartel and Wang suggest that researchers could design guide RNAs with weak centers, which would require more perfect pairing in the tail end, so that the guide RNA will be less likely to bind non-target RNAs; only the perfect pairing of the target’s RNA sequence would suffice.
Altogether, Wang and Bartel’s findings explain how small differences between guide RNAs can make such large differences in the efficacy of RNAi, providing a rationale for the long-standing RNAi design guidelines. Some of the findings even suggest new guidelines that could help with future synthetic guide RNA designs.
“Discovering the interplay between the center and tail end of the guide RNA was unexpected and satisfying,” says Bartel, who is also a professor at the Massachusetts Institute of Technology and a Howard Hughes Medical Investigator. “It explains why, even though the guidelines suggested that tail-end sequence doesn’t matter, the target RNAs that are sliced in our cells do have pairing to the tail end. This observation could prove useful to reduce off-target effects in RNAi therapeutics.”
Jonathan Weissman is the senior author on a recent study on silencing a prion protein's expression. Prions cause devastating neurodegenerative disorders such as dementia, Huntington's, Parkinson's, and Lou Gehrig's disease. Silencing genes represents a step towards a therapeutic model for treating these diseases in humans.
Karen Weintraub | USA TODAY
June 27, 2024
CAMBRIDGE, Mass. ‒ Sonia Vallabh watched helplessly as her 51-year-old mother rapidly descended into dementia and died. It didn’t take long for Vallabh to realize she was destined for the same rare genetic fate.
Vallabh and her husband did what anyone would want to do in their situation: They decided to fight.
Armed with little more than incredible intellect and determination they set out to conquer her destiny.
A dozen years later, they’ve taken a major step in that direction, finding a way to shut off enough genetic signals to hold off the disease.
And in the process of trying to rescue Vallabh, they may save many, many others as well.
The same approach should also work against diseases such as Huntington’s, Parkinson’s, Lou Gehrig’s disease and even Alzheimer’s, which result from the accumulation of toxic proteins. If it works as well as they think, it could also be useful against a vast array of other diseases that can be treated by shutting off genes.
“It doesn’t have to be the brain. It could be the muscles. It could be the kidneys. It could be really anywhere in the body where we have not easily been able to do these things before,” said Dr. Kiran Musunuru, a cardiologist and geneticist at the University of Pennsylvania’s Perelman School of Medicine, who wasn’t involved in the research but wrote a perspective accompanying the paper.
So far, they’ve proven it only in mice.
“The data are good as far as they go,” Vallabh said this week from her office at the Broad Institute of Harvard and the Massachusetts Institute of Technology, where she has worked since getting a Ph.D. at Harvard. She had already gotten a law degree from the university, but she and Minikel, then a transportation planner, both pursued biology degrees after her mother’s death. Now, they work together at the Broad.
“We’re far from this being a drug,” Vallabh said. “There’s always, always reason for caution. Sadly, everything is always more likely to fail than succeed.
“But there is justifiable reason for optimism.”
A terrible disease
The disease that killed Vallabh’s mother was one of a group of conditions called prion diseases. These include mad cow disease, which affects mostly cattle, scrapie, which affects sheep, and Creutzfeldt-Jakob disease, which kills about 350 Americans a year ‒ most within months of their first symptom.
These diseases are triggered when the prion protein found in all normal brains starts misfolding for some reason, as yet unknown.
“Prion disease can strike anybody,” Vallabh said, noting the 1 in 6,000 risk to the general population.
Though prion diseases are, in some cases, contagious, a federal study earlier this year concluded that chronic wasting disease, found in deer, elk and moose, is very unlikely to pass to people who eat the meat of sick animals.
In Vallabh’s case, the cause is genetic. Vallabh discovered after her mother’s death that she carries the same variant of the same gene that caused her mother’s disease, meaning she will certainly develop it.
The only question is when.
“The age of onset is extremely unpredictable,” Vallabh said. “Your parent’s age of onset doesn’t actually predict anything.”
How the gene-editing tool works
Vallabh and Minikel approached colleagues at the Whitehead Institute a biomedical research institute next to the Broad. They asked to collaborate on a new gene-editing approach to turn off Vallabh’s disease gene. The technique developed by Whitehead scientists is called CHARM (for Coupled Histone tail Autoinhibition Release of Methyltransferase).
While previous gene-editing tools have been described as scissors or erasers, Musunuru described CHARM as volume control, allowing scientists to tune a gene up or down. It has three advantages over previous strategies, he said.
The device is tiny, so it fits easily inside the virus needed to deliver it. Other gene-editing tools, like CRISPR, are bigger, which means they need to be broken into pieces and much more of the virus is needed to deliver those pieces to the brain, risking a dangerous immune reaction.
CHARM, Musunuru said, is “easier to deliver to hard-to-deliver spaces like the brain.”
At least in the mouse, it also seems to have reached throughout the brain, making the desired genetic change without other, unwanted ones, Musunuru said.
And finally, the research team figured out a way to turn the gene editor off after its work was done. “If it’s sticking around, there’s the potential for genetic mischief,” Musunuru said.
One shot on goal
While researchers, including Vallabh, continue to work to perfect an approach, the clock for Vallabh and others is ticking.
Right now there’s no viable treatment and if it takes too long to develop one, Vallabh will miss her window. Once the disease process starts, like a runaway train, it’ll be much harder to stop than it would be to just shut the gene off in the first place.
The more prion protein in the brain, the more likely it is to misfold. And the more likely it is for the disease to spread, a process that co-opts the natural form of the protein and converts it to the toxic form.
That’s why getting rid of as much of it as possible makes sense, said Jonathan Weissman, the senior author of the study, who leads a Whitehead lab.
“The biology is really clear. The need (for a cure) is so compelling,” Weissman said.
Every cell in the brain has the gene for making the prion protein. By silencing even 50% of those genes, Weissman figures he can prevent the disease. In mice, CHARM silenced up to 80% to 90%.
“We’ve figured out what to deliver. Now we have to figure out how to deliver it,” he said.
Another of the paper’s co-authors, the Broad’s Ben Deverman, published a study late last year showing he could deliver a gene-therapy-carrying virus throughout the brain. Others are developing other viral delivery systems.
Vallabh and Minikel have hedged their bets, helping to develop a so-called antisense oligonucleotide, or ASO, which uses another path for stopping the gene from making the prion protein.
The ASO, which is in early trials in people by a company called Ionis Pharmaceuticals, requires regular treatment rather than the one-and-done of gene therapy. Recruitment for that trial had to be paused in April because the number of would-be volunteers outstripped the available slots.
Vallabh isn’t ready yet to start any treatment yet herself.
“She has one shot on goal,” Musunuru said. “At some point, she’ll have to decide what’s the best strategy.”
In the meantime, the clock Vallabh can’t see continues to tick toward the onset.
She and Minikel stay exceedingly busy with their research along with their daughter, almost 7, and 4-year-old son ‒ both born via IVF and preimplantation genetic testing to ensure they wouldn’t inherit her genetic curse. (They were super lucky, Vallabh notes, to be living in Massachusetts where IVF is at least “approachable” financially.)
“There is a mountain ahead of us,” Vallabh said of the path to a cure. “There’s still a lot of hurdles, there’s still a lot to figure out.”
A new gene-silencing tool shows promise as a future therapy against prion diseases and paves the way for new approaches to treating disease.
Greta Friar | Whitehead Institute
June 27, 2024
Drug development is typically slow: The pipeline from basic research discoveries that provide the basis for a new drug to clinical trials and then production of a widely available medicine can take decades. But decades can feel impossibly far off to someone who currently has a fatal disease. Broad Institute of MIT and Harvard Senior Group Leader Sonia Vallabh is acutely aware of that race against time, because the topic of her research is a neurodegenerative and ultimately fatal disease — fatal familial insomnia, a type of prion disease — that she will almost certainly develop as she ages.
Vallabh and her husband, Eric Minikel, switched careers and became researchers after they learned that Vallabh carries a disease-causing version of the prion protein gene and that there is no effective therapy for fatal prion diseases. The two now run a lab at the Broad Institute, where they are working to develop drugs that can prevent and treat these diseases, and their deadline for success is not based on grant cycles or academic expectations but on the ticking time bomb in Vallabh’s genetic code.
That is why Vallabh was excited to discover, when she entered into a collaboration with Whitehead Institute for Biomedical Research member Jonathan Weissman, that Weissman’s group likes to work at full throttle. In less than two years, Weissman, Vallabh, and their collaborators have developed a set of molecular tools called CHARMs that can turn off disease-causing genes such as the prion protein gene — as well as, potentially, genes coding for many other proteins implicated in neurodegenerative and other diseases — and they are refining those tools to be good candidates for use in human patients. Although the tools still have many hurdles to pass before the researchers will know if they work as therapeutics, the team is encouraged by the speed with which they have developed the technology thus far.
“The spirit of the collaboration since the beginning has been that there was no waiting on formality,” Vallabh says. “As soon as we realized our mutual excitement to do this, everything was off to the races.”
Co-corresponding authors Weissman and Vallabh and co-first authors Edwin Neumann, a graduate student in Weissman’s lab, and Tessa Bertozzi, a postdoc in Weissman’s lab, describe CHARM — which stands for Coupled Histone tail for Autoinhibition Release of Methyltransferase — in a paper published today in the journal Science.
“With the Whitehead and Broad Institutes right next door to each other, I don’t think there’s any better place than this for a group of motivated people to move quickly and flexibly in the pursuit of academic science and medical technology,” says Weissman, who is also a professor of biology at MIT and a Howard Hughes Medical Institute Investigator. “CHARMs are an elegant solution to the problem of silencing disease genes, and they have the potential to have an important position in the future of genetic medicines.”
To treat a genetic disease, target the gene
Prion disease, which leads to swift neurodegeneration and death, is caused by the presence of misshapen versions of the prion protein. These cause a cascade effect in the brain: the faulty prion proteins deform other proteins, and together these proteins not only stop functioning properly but also form toxic aggregates that kill neurons. The most famous type of prion disease, known colloquially as mad cow disease, is infectious, but other forms of prion disease can occur spontaneously or be caused by faulty prion protein genes.
Most conventional drugs work by targeting a protein. CHARMs, however, work further upstream, turning off the gene that codes for the faulty protein so that the protein never gets made in the first place. CHARMs do this by epigenetic editing, in which a chemical tag gets added to DNA in order to turn off or silence a target gene. Unlike gene editing, epigenetic editing does not modify the underlying DNA — the gene itself remains intact. However, like gene editing, epigenetic editing is stable, meaning that a gene switched off by CHARM should remain off. This would mean patients would only have to take CHARM once, as opposed to protein-targeting medications that must be taken regularly as the cells’ protein levels replenish.
Research in animals suggests that the prion protein isn’t necessary in a healthy adult, and that in cases of disease, removing the protein improves or even eliminates disease symptoms. In a person who hasn’t yet developed symptoms, removing the protein should prevent disease altogether. In other words, epigenetic editing could be an effective approach for treating genetic diseases such as inherited prion diseases. The challenge is creating a new type of therapy.
Fortunately, the team had a good template for CHARM: a research tool called CRISPRoff that Weissman’s group previously developed for silencing genes. CRISPRoff uses building blocks from CRISPR gene editing technology, including the guide protein Cas9 that directs the tool to the target gene. CRISPRoff silences the targeted gene by adding methyl groups, chemical tags that prevent the gene from being transcribed, or read into RNA, and so from being expressed as protein. When the researchers tested CRISPRoff’s ability to silence the prion protein gene, they found that it was effective and stable.
Several of its properties, though, prevented CRISPRoff from being a good candidate for a therapy. The researchers’ goal was to create a tool based on CRISPRoff that was just as potent but also safe for use in humans, small enough to deliver to the brain, and designed to minimize the risk of silencing the wrong genes or causing side effects.
From research tool to drug candidate
Led by Neumann and Bertozzi, the researchers began engineering and applying their new epigenome editor. The first problem that they had to tackle was size, because the editor needs to be small enough to be packaged and delivered to specific cells in the body. Delivering genes into the human brain is challenging; many clinical trials have used adeno-associated viruses (AAVs) as gene-delivery vehicles, but these are small and can only contain a small amount of genetic code. CRISPRoff is way too big; the code for Cas9 alone takes up most of the available space.
The Weissman lab researchers decided to replace Cas9 with a much smaller zinc finger protein (ZFP). Like Cas9, ZFPs can serve as guide proteins to direct the tool to a target site in DNA. ZFPs are also common in human cells, meaning they are less likely to trigger an immune response against themselves than the bacterial Cas9.
Next, the researchers had to design the part of the tool that would silence the prion protein gene. At first, they used part of a methyltransferase, a molecule that adds methyl groups to DNA, called DNMT3A. However, in the particular configuration needed for the tool, the molecule was toxic to the cell. The researchers focused on a different solution: Instead of delivering outside DNMT3A as part of the therapy, the tool is able to recruit the cell’s own DNMT3A to the prion protein gene. This freed up precious space inside of the AAV vector and prevented toxicity.
The researchers also needed to activate DNMT3A. In the cell, DNMT3A is usually inactive until it interacts with certain partner molecules. This default inactivity prevents accidental methylation of genes that need to remain turned on. Neumann came up with an ingenious way around this by combining sections of DNMT3A’s partner molecules and connecting these to ZFPs that bring them to the prion protein gene. When the cell’s DNMT3A comes across this combination of parts, it activates, silencing the gene.
“From the perspectives of both toxicity and size, it made sense to recruit the machinery that the cell already has; it was a much simpler, more elegant solution,” Neumann says. “Cells are already using methyltransferases all of the time, and we’re essentially just tricking them into turning off a gene that they would normally leave turned on.”
Testing in mice showed that ZFP-guided CHARMs could eliminate more than 80 percent of the prion protein in the brain, while previous research has shown that as little as 21 percent elimination can improve symptoms.
Once the researchers knew that they had a potent gene silencer, they turned to the problem of off-target effects. The genetic code for a CHARM that gets delivered to a cell will keep producing copies of the CHARM indefinitely. However, after the prion protein gene is switched off, there is no benefit to this, only more time for side effects to develop, so they tweaked the tool so that after it turns off the prion protein gene, it then turns itself off.
Meanwhile, a complementary project from Broad Institute scientist and collaborator Benjamin Deverman’s lab, focused on brain-wide gene delivery and published in Science on May 17, has brought the CHARM technology one step closer to being ready for clinical trials. Although naturally occurring types of AAV have been used for gene therapy in humans before, they do not enter the adult brain efficiently, making it impossible to treat a whole-brain disease like prion disease. Tackling the delivery problem, Deverman’s group has designed an AAV vector that can get into the brain more efficiently by leveraging a pathway that naturally shuttles iron into the brain. Engineered vectors like this one make a therapy like CHARM one step closer to reality.
Thanks to these creative solutions, the researchers now have a highly effective epigenetic editor that is small enough to deliver to the brain, and that appears in cell culture and animal testing to have low toxicity and limited off-target effects.
“It’s been a privilege to be part of this; it’s pretty rare to go from basic research to therapeutic application in such a short amount of time,” Bertozzi says. “I think the key was forming a collaboration that took advantage of the Weissman lab’s tool-building experience, the Vallabh and Minikel lab’s deep knowledge of the disease, and the Deverman lab’s expertise in gene delivery.”
Looking ahead
With the major elements of the CHARM technology solved, the team is now fine-tuning their tool to make it more effective, safer, and easier to produce at scale, as will be necessary for clinical trials. They have already made the tool modular, so that its various pieces can be swapped out and future CHARMs won’t have to be programmed from scratch. CHARMs are also currently being tested as therapeutics in mice.
The path from basic research to clinical trials is a long and winding one, and the researchers know that CHARMs still have a way to go before they might become a viable medical option for people with prion diseases, including Vallabh, or other diseases with similar genetic components. However, with a strong therapy design and promising laboratory results in hand, the researchers have good reason to be hopeful. They continue to work at full throttle, intent on developing their technology so that it can save patients’ lives not someday, but as soon as possible.