

To create proteins, DNA is transcribed into RNA, and that RNA is then “translated” into protein. Between the creation of the RNA and the translation to protein is often a step called splicing. During splicing, segments called introns are removed, and the remaining pieces, called exons, are joined together to form the blueprint for translation. By splicing together different exons, the cell can create different proteins from the same section of genetic code. When splicing goes awry, it can lead to diseases and cancers.
New research recently published in Disease Models & Mechanisms from the Calo Lab in the Department of Biology at MIT has identified the mechanism for how cells respond to disruptions in splicing, which involves activating a cellular stress response. The stress response, once activated, causes widespread effects, including changes to cell metabolism.
Researchers have discovered cellular stress responses for other core cellular processes, such as ribosome biogenesis. However, this is the first time researchers have identified how cells respond to perturbing the splicing process.
A particular protein acts as a kind of canary in a coal mine: Mdm2, which responds to a broad range of splicing disruptions. Mdm2 does not cause a stress response by itself. Rather, Mdm2 is itself spliced differently in response to splicing disruptions. Downstream, the alternative splicing of Mdm2 leads to the activation of a protein called p53, which is known to orchestrate a cascade of responses to stress.
Researchers have long wondered why some cell types seem more sensitive to splicing disruptions than others. For example, some disorders caused by mutations in proteins that perform RNA splicing, despite affecting the whole organism, induce more noticeable changes in tissues derived from the neural crest—a collection of stem cells that contributes to the formation of the face, jaw, retinas, limbs, and heart during development. Certain splicing inhibitors have also increased the effectiveness of some cancer treatments, but the mechanism is unknown.
One of the p53-induced stress responses includes changing the metabolism of cells and how they use sugars, which may explain why some cells are more sensitive to splicing disruptions than others. Inhibiting glycolysis, the reactions that extract energy from glucose, can affect how cells divide and migrate.
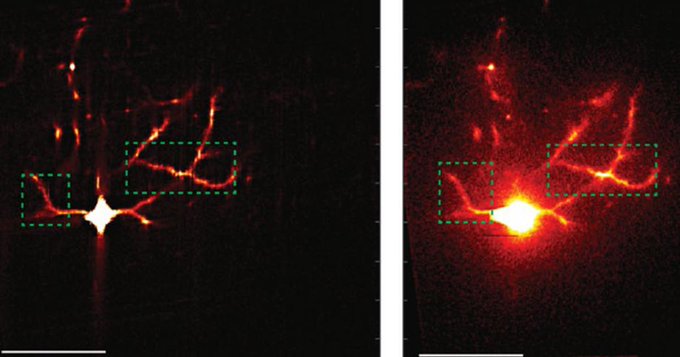
The way cells divide and migrate is critical during development; in experiments, zebrafish treated with glycolysis inhibitors exhibited similar changes to craniofacial features as those where splicing was disrupted. Cancerous cells, too, are known to require high levels of sugar metabolism and, therefore, may be especially sensitive to treatments that induce changes in the splicing pathway.
The researchers knocked down genes to mimic milder splicing disruptions instead of knocking them out entirely. Splicing is so essential that knocking out the splicing machinery can lead to extreme responses like cell death. In organismal models like zebrafish, those severe phenotypes don’t accurately reflect how splicing disruptions present in human diseases.
First author Jade Varineau, a graduate student in the Calo lab, was drawn to the project because it allowed her to explore what was happening at the RNA and cellular level while also observing how splicing perturbations were affecting the whole organism.
“I think this data can help us reframe the way we think about diseases and cancers that are impacted by splicing—that a treatment that works for one may work for another because all the symptoms may stem from the same cellular response,” Varineau says.
Although the results indicate how cells broadly respond to splicing perturbations, the mechanism for how disruptions in splicing induce the alternate splicing of Mdm2 remains unclear. Senior author Eliezer Calo says the lab is also exploring how splicing mechanisms may be altered for things like cancer. Their work, he says, opens the door for further exploration of cell-type specificity of genetic disorders and improvements in cancer treatments using splicing inhibitors.
“We know that the sensor is encoded in the gene Mdm2—what are the molecules that allow Mdm2 to act as a sensor, and how does the sensor malfunction for things like cancer?” Calo says. “The next step is to find out how the sensor works.”
One of the immune system’s primary roles is to detect and kill cells that have acquired cancerous mutations. However, some early-stage cancer cells manage to evade this surveillance and develop into more advanced tumors.
A new study from MIT and Dana-Farber Cancer Institute has identified one strategy that helps these precancerous cells avoid immune detection. The researchers found that early in colon cancer development, cells that turn on a gene called SOX17 can become essentially invisible to the immune system.
If scientists could find a way to block SOX17 function or the pathway that it activates, this may offer a new way to treat early-stage cancers before they grow into larger tumors, the researchers say.
“Activation of the SOX17 program in the earliest innings of colorectal cancer formation is a critical step that shields precancerous cells from the immune system. If we can inhibit the SOX17 program, we might be better able to prevent colon cancer, particularly in patients that are prone to developing colon polyps,” says Omer Yilmaz, an MIT associate professor of biology, a member of MIT’s Koch Institute for Integrative Cancer Research, and one of the senior authors of the study.
Judith Agudo, a principal investigator at Dana-Farber Cancer Institute and an assistant professor at Harvard Medical School, is also a senior author of the study, which appears today in Nature. The paper’s lead author is MIT Research Scientist Norihiro Goto. Other collaborators include Tyler Jacks, a professor of biology and a member of MIT’s Koch Institute; Peter Westcott, a former Jacks lab postdoc who is now an assistant professor at Cold Spring Harbor Laboratory; and Saori Goto, an MIT postdoc in the Yilmaz lab.
Immune evasion
Colon cancer usually arises in long-lived cells called intestinal stem cells, whose job is to continually regenerate the lining of the intestines. Over their long lifetime, these cells can accumulate cancerous mutations that lead to the formation of polyps, a type of premalignant growth that can eventually become metastatic colon cancer.
To learn more about how these precancerous growths evade the immune system, the researchers used a technique they had previously developed for growing mini colon tumors in a lab dish and then implanting them into mice. In this case, the researchers engineered the tumors to express mutated versions of cancer-linked genes Kras, p53, and APC, which are often found in human colon cancers.
Once these tumors were implanted in mice, the researchers observed a dramatic increase in the tumors’ expression of SOX17. This gene encodes a transcription factor that is normally active only during embryonic development, when it helps to control development of the intestines and the formation of blood vessels.
The researchers’ experiments revealed that when SOX17 is turned on in cancer cells, it helps the cells to create an immunosuppressive environment. Among its effects, SOX17 prevents cells from synthesizing the receptor that normally detects interferon gamma, a molecule that is one of the immune system’s primary weapons against cancer cells.
Without those interferon gamma receptors, cancerous and precancerous cells can simply ignore messages from the immune system, which would normally direct them to undergo programmed cell death.
“One of SOX17’s main roles is to turn off the interferon gamma signaling pathway in colorectal cancer cells and in precancerous adenoma cells. By turning off interferon gamma receptor signaling in the tumor cells, the tumor cells become hidden from T cells and can grow in the presence of an immune system,” Yilmaz says.
Without interferon gamma signaling, cancer cells also minimize their production of molecules called MHC proteins, which are responsible for displaying cancerous antigens to the immune system. The cells’ insensitivity to interferon gamma also prevents them from producing immune molecules called chemokines, which normally recruit T cells that would help destroy the cancerous cells.
Targeting SOX17
When the researchers generated colon tumor organoids with SOX17 knocked out, and implanted those into mice, the immune system was able to attack those tumors much more effectively. This suggests that preventing cancer cells from turning off SOX17 could offer a way to treat colon cancer in its earliest stages.
“Just by turning off SOX17 in fairly complex tumors, we were able to essentially obliterate the ability of these tumor cells to persist,” Goto says.
As part of their study, the researchers also analyzed gene expression data from patients with colon cancer and found that SOX17 tended to be highly expressed in early-stage colon cancers but dropped off as the tumors became more invasive and metastatic.
“We think this makes a lot of sense because as colorectal cancers become more invasive and metastatic, there are other mechanisms that create an immunosuppressive environment,” Yilmaz says. “As the colon cancer becomes more aggressive and activates these other mechanisms, then there’s less importance for SOX17.”
Transcription factors such as SOX17 are considered difficult to target using drugs, in part because of their disorganized structure, so the researchers now plan to identify other proteins that SOX17 interacts with, in hopes that it might be easier to block some of those interactions.
The researchers also plan to investigate what triggers SOX17 to turn on in precancerous cells.
The research was funded by the MIT Stem Cell Initiative via Fondation MIT, the National Institutes of Health/National Cancer Institute, and a Koch Institute-Dana Farber Harvard Cancer Center Bridge Project grant.

When Albert E. Almada PhD ’13 embarks on a new project, he always considers two criteria instilled in him during his time as a graduate student in the Department of Biology at MIT.
“If you want to make a big discovery, you have to approach it from a unique perspective — a unique angle,” Almada says. “You also have to be willing to dive into the unknown and go to the leading edge of your field.”
This is not without its challenges — but with an innovative spirit, Almada says, one can find ways to apply technologies and approaches to a new area of research where a roadmap doesn’t yet exist.
Now an assistant professor of orthopedic surgery and stem cell biology and regenerative medicine at the Keck School of Medicine of the University of Southern California (USC), Almada studies the mechanics of how stem cells rebuild tissues after trauma and how stem cell principles are dysregulated and drive conditions like degenerative disease and aging, exploring these topics through an evolutionary lens.
He’s also trying to solve a mystery that has intrigued scientists for centuries: Why can some vertebrate species like fish, salamanders, and lizards regenerate entire body parts, but mammals cannot? Almada’s laboratory at USC tackles these critical questions in the musculoskeletal system.
Almada’s fascination with muscle development and regeneration can be traced back to growing up in southern California. Almada’s brother had a degenerative muscle disease called Duchenne muscular dystrophy — and, while Almada grew stronger and stronger, his brother grew weaker and weaker. Last summer, Almada’s brother, unfortunately, lost his battle with his disorder at the age of 41.
“Watching his disease progress in those early years is what inspired me to become a scientist,” Almada recalls. “Sometimes science can be personal.”
Almada went to the University of California at Irvine for his undergraduate degree, majoring in biological sciences. During his summers, he participated in the Undergraduate Research Program (URP) at the Cold Spring Harbor Laboratory and the MIT Summer Research Program-Bio (now the Bernard S. and Sophie G. Gould MIT Summer Research Program in Biology, BSG-MSRP-Bio), where he saw the passion, rigor, and drive that solidified his desire to pursue a PhD.
Despite his interest in clinical applications, skeletal muscle, and regenerative biology, Almada was drawn to the Department of Biology at MIT, which is focused on basic fundamental research.
“I was willing to bet that it all came down to understanding basic cellular processes and things going wrong with the cell and how it interacts with its environment,” he says. “The MIT biology program really helped me define an identity for myself and gave me a template for how to tackle clinical problems from a molecular perspective.”
Almada’s PhD thesis work was based on a curious finding that Phillip Sharp, Institute Professor emeritus, professor emeritus of biology, and intramural faculty at the Koch Institute for Integrative Cancer Research, had made in 2007 — that transcription, the process of copying DNA into a messenger molecule called RNA, can occur in both directions at gene promoters. In one direction, it was long understood that fully formed mRNA is transcribed and can be used as a blueprint to make a protein. The transcription Sharp observed, in the opposite direction, results in a very short RNA that is not used as a gene product blueprint.
Almada’s project dug into what those short RNA molecules are — their structure and sequence, and why they’re not produced the same way that coding messenger RNA is. In two papers published in PNAS and Nature, Almada and colleagues discovered that a balance between splicing and transcription termination signals controls the length of an RNA. This finding has wider implications because toxic RNAs are produced and can build up in several degenerative diseases; being able to splice out or shorten RNAs to remove the harmful segments could be a potential therapeutic treatment.
“That experience convinced me that if I want to make big discoveries, I have to focus on basic science,” he says. “It also gave me the confidence that if I can succeed at MIT, I can succeed just about anywhere and in any field of biology.”
At the time Almada was in graduate school, there was a lot of excitement about transcription factor reprogramming. Transcription factors are the proteins responsible for turning on essential genes that tell a cell what to be and how to behave; a subset of them can even theoretically turn one cell type into another.
Almada began to wonder whether a specialized set of transcription factors instructs stem cells to rebuild tissues after trauma. After MIT, Almada moved on to a postdoctoral position in the lab of Amy Wagers, a leader in muscle stem cell biology at Harvard University, to immerse himself in this problem.
In many tissues in our bodies, a population of stem cells typically exists in an inactive, non-dividing state called quiescence. Once activated, these stem cells interact with their environment, sense damage signals, and turn on programs of proliferation and differentiation, as well as self-renewal, which is critical to maintaining a pool of stem cells in the tissue.
One of the biggest mysteries in the field of regenerative biology is how stem cells transition from dormancy into that activated, highly regenerative state. The body’s ability to turn on stem cells, including those in the skeletal muscle system, declines as we age and is often dysregulated in degenerative diseases — diseases like the one Almada’s brother suffered from.
In a study Almada published in Cell Reports several years ago, he identified a family of transcription factors that work together to turn on a critical regenerative gene program within hours of muscle trauma. This program drives muscle stem cells out of quiescence and speeds up healing.
“Now my lab is studying this regenerative program and its potential dysregulation in aging and degenerative muscle diseases using mouse and human models,” Almada says. “We’re also drawing parallels with super-healing species like salamanders and lizards.”
Recently, Almada has been working on characterizing the molecular and functional properties of stem cells in lizards, attempting to understand how the genes and pathways differ from mammalian stem cells. Lizards can regenerate massive amounts of skeletal muscle from scratch — imagine if human muscle tissue could be regrown as seamlessly as a lizard’s tail can. He is also exploring whether the tail is unique, or if stem cells in other tissues in lizards can regenerate faster and better than the tail, by comparing analogous injuries in a mouse model.
“This is a good example of approaching a problem from a new perspective: We believe we’re going to discover new biology in lizards that we can use to enhance skeletal muscle growth in vulnerable human populations, including those that suffer from deadly muscle disorders,” Almada says.
In just three years of starting his faculty position at USC, his work and approach have already received recognition in academia, with junior faculty awards from the Baxter Foundation and the Glenn Foundation/American Federation of Aging Research. He also received his first RO1 award from the National Institutes of Health with nearly $3 million in funding. Almada and his first graduate student, Alma Zuniga Munoz, were also awarded the HHMI Gilliam Fellowship last summer. Zuniga Munoz is the first to be recognized with this award at USC; fellowship recipients, student and advisor pairs, are selected with the goal of preparing students from underrepresented groups for leadership roles in science.
Almada himself is a second-generation Mexican American and has been involved in mentoring and training throughout his academic career. He was a graduate resident tutor for Spanish House at MIT and currently serves as the chair of the Diversity, Equity, and Inclusion Committee in the Department of Stem Cell Biology and Regenerative Medicine at USC; more than half of his lab members identify as members of the Hispanic community.
“The focus has to be on developing good scientists,” Almada says. “I learned from my past research mentors the importance of putting the needs of your students first and providing a supportive environment for everyone to excel, no matter where they start.”
As a mentor and researcher, Almada knows that no question and no challenge is off limits — foundations he built in Cambridge, where his graduate studies focused on teaching him to think, not just do.
“Digging deep into the science is what MIT taught me,” he says. “I’m now taking all of my knowledge in molecular biology and applying it to translationally oriented questions that I hope will benefit human health and longevity.”
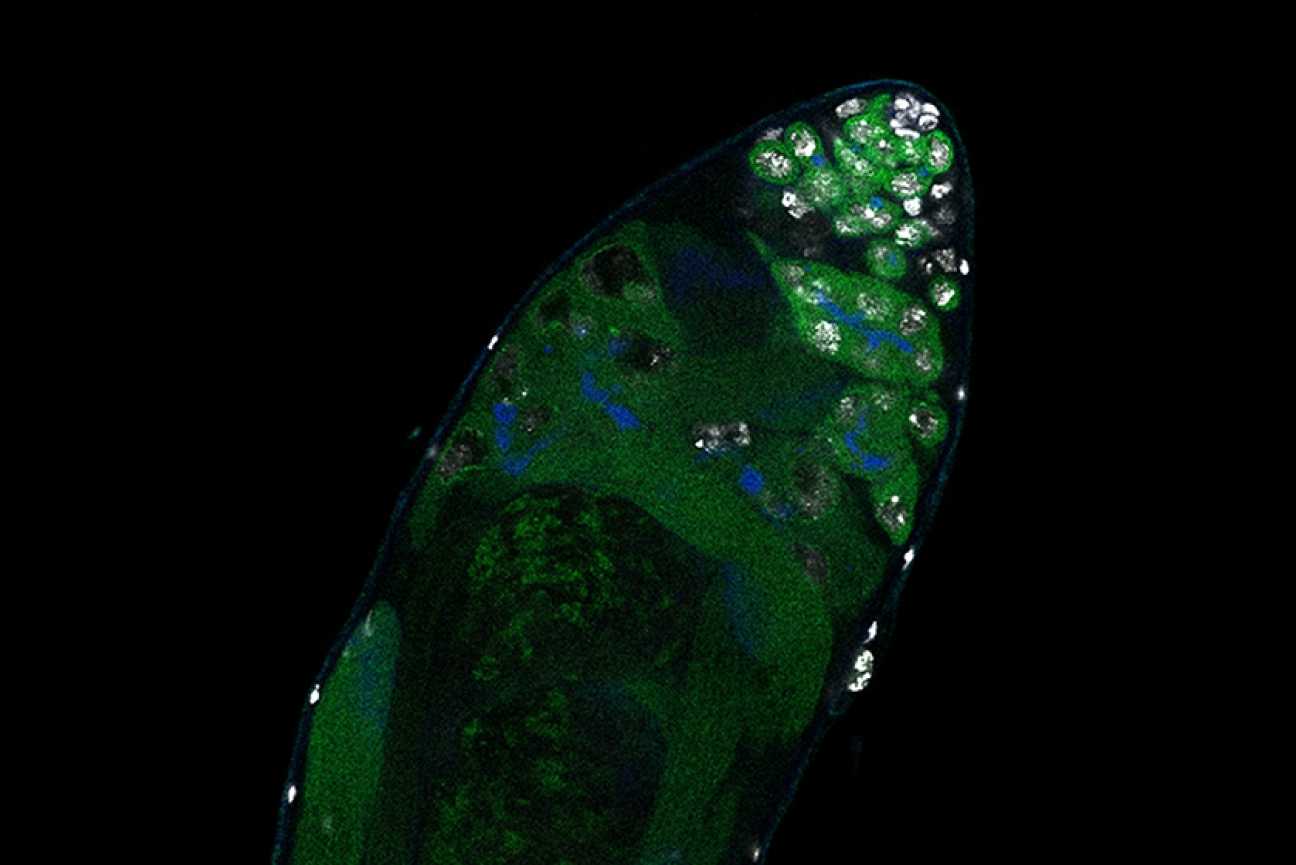
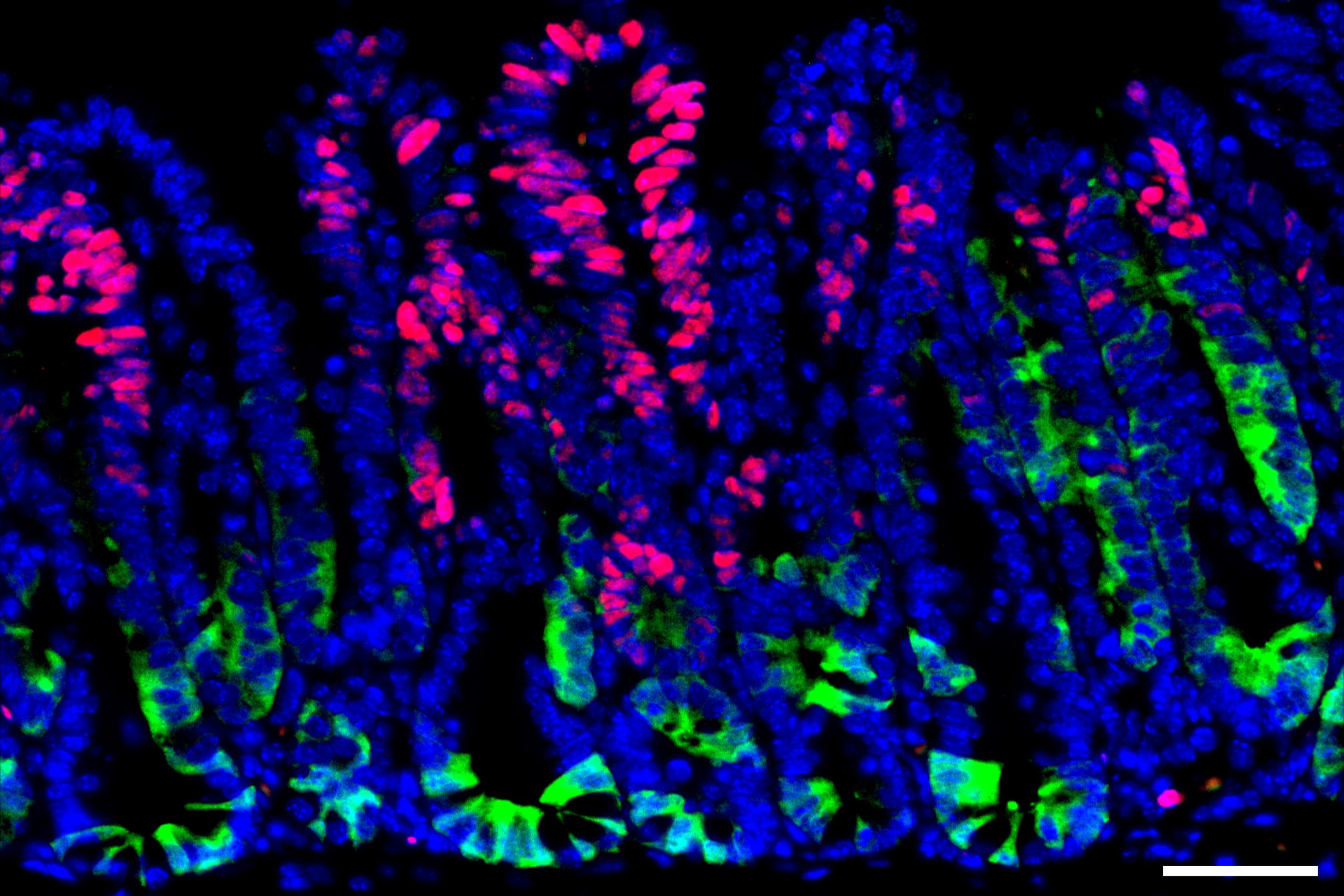
Germline stem cells are the pool of stem cells capable of becoming eggs or sperm. They divide asymmetrically, such that one of the cells resulting from a division is another stem cell and the other is a differentiated cell, which has progressed one step further down the path towards becoming an egg or sperm. Researchers have thought that this asymmetrical division served to replenish the pool of stem cells—making sperm or eggs, but also making more stem cells to produce future sperm or eggs. However, the germline has another way to replenish itself: cells that have differentiated only one or two steps down the path to becoming eggs or sperm are capable of reverting into stem cells. Why, then, do stem cells divide asymmetrically?
New research from Whitehead Institute Member Yukiko Yamashita, who is also a professor of biology at the Massachusetts Institute of Technology and an HHMI Investigator, and former postdoc in her lab Jonathan Nelson shows that asymmetrical division in germline stem cells serves a different but equally important purpose in male fruit flies (Drosophila melanogaster), a common model animal for germline research. The work, published in the journal Proceedings of the National Academy of Sciences (PNAS) on November 13, suggests that in flies, germline stem cells divide asymmetrically in order to unequally split a certain kind of DNA, called ribosomal DNA (rDNA), between the two dividing cells and then keep the cell with more rDNA in the stem cell pool. This is necessary in order to keep the germline viable over generations of cell divisions, and so to keep individual flies fertile and capable of reproduction. The researchers show that only germline stem cells, and not other types of germ cells, drive this process, and explain why stem cells’ asymmetric divisions make them uniquely suited to maintaining rDNA.
Ribosomal DNA is critical to maintain in the germline because it contains the instructions for making a major part of ribosomes, the cellular machines that build proteins from genetic instructions. Proteins are the main workhorses of the cell, and so cells need to make many ribosomes in order to build all of the proteins that they need. Consequently, rDNA exists as many copies repeated in a row of the code for components of the ribosome. All of these repeats make it easy for the cell to mass produce ribosomes, but they also come with a risk: repetitive DNA is prone to losing repeats during cell division. When the cell’s rDNA is copied, it’s easy for a few of the many identical repeats to get cut out, so that the resulting copy of the genome has fewer rDNA repeats than the original.
Most cells can afford to lose a few rDNA repeats without too many negative effects, but the germline cannot. Whereas other cells die with the body they are in, germ cells produce eggs and sperm that will form a new body, which produces new germ cells, and so on. The germ cell lineage is effectively immortal. Over the course of its endless cycle of cell division, the loss of rDNA repeats would add up until the cells became dysfunctional and then died. This would make the individual bearing those germ cells infertile, and so cause their lineage to go extinct.
Researchers have known that germ cells have some way to regain rDNA repeats when the number gets too low—if germ cells couldn’t do this, none of us would exist—but the details of how cells achieve this have been largely mysterious. One proposed model was that when a germ cell divides, sometimes it might divide up its rDNA unequally between the two resulting cells, so that one cell would gain rDNA repeats. Yamashita and Nelson have previously found evidence that this model is correct, and they discovered some of the specific mechanisms that enable it to happen. In a 2023 PNAS paper, the researchers showed that a retrotransposon, a “selfish” genetic element whose function is to make more copies of itself, actually helps germ cells maintain rDNA. During cell division, the retrotransposon R2 slices open one copy of the chromosome containing rDNA in its quest to insert extra copies of itself into the genome. The cell tries to repair the break using the copy on the other intact chromosome, but the tricky nature of repetitive DNA can cause the cell to lose its place, so that it stitches a stretch of rDNA repeats from one copy of the chromosome into the other copy instead.
Through this process, the germline can boost the level of rDNA in a cell—but only by as much as another cell loses. How does this win-lose exchange lead to an overall increase in rDNA levels across the germline cell population to compensate for lost rDNA? In this latest work, Yamashita and Nelson show through mathematical modeling that in cells that divide symmetrically, it would not. Gains and losses in rDNA through this form of exchange would occur essentially at random and cancel each other out over time.
Now consider an asymmetric division. After a germline stem cell divides, the cell that differentiates will go through a few more divisions and ultimately create a specific number of sperm cells–the number happens to be sixty-four. If this daughter cell gets the chromosome with more rDNA repeats, then that would lead to sixty-four sperm with more rDNA repeats—but that would be it, as the sperm have exited the pool of replicating germline stem cells.
However, the daughter cell that remains a germline stem cell will divide again to create a differentiated cell (which will become sixty-four sperm) and another stem cell, which will divide again, leading to another sixty-four sperm and another stem cell—and so on. All of these cells, including many sperm, would inherit the higher number of rDNA repeats. Furthermore, at each division, there would be an opportunity for another unequal split of rDNA. As long as the stem cell always gets the boost in rDNA, then the cumulative number of rDNA repeats would keep growing in the overall population over time—and Yamashita and colleagues’ past work shows that the germline can ensure this. A 2022 Science Advances paper from Yamashita and then-postdoc in her lab George Watase showed that when a germline stem cell divides, the DNA strand with more rDNA repeats is tagged with a protein that the researchers named Indra, which helps mark it to stay in the daughter cell that will become another stem cell. Yamashita and Nelson’s new paper includes mathematical modeling by second author Tomohiro Kumon, a postdoc in Yamashita’s lab, that proves that this is not only sufficient to restore the level of rDNA repeats over time, but that it is the most effective and efficient way for the germline to do so.
“There was this problem with the unequal exchange model of rescuing rDNA, because every cell that gained rDNA did so at the expense of another that was losing it,” Nelson says. “What we show here is that the reason why there’s a bias towards gain in the germline is because this process is happening within these asymmetrically dividing germline stem cells that can gain and gain and gain, while the cells that lose rDNA exit the cycle and so have a limited effect.”
The researchers complemented their mathematical modeling with evidence that the process to increase rDNA repeats occurs primarily or solely in germline stem cells. They found that when the number of rDNA repeats got low enough, then expression of R2 and the presence of double-stranded DNA breaks both increased in germline stem cells, but not significantly in other germ cell types.
Yamashita and Nelson propose that the different cell types in the germline take on different functions to create a pipeline for maximizing the health of future sperm. Germ cells that are one or two steps down the path of differentiation from stem cells are essentially identical to them, to the point that they can be difficult to tell apart in testing, but they divide symmetrically. They are also much more sensitive to DNA damage; the researchers found that R2 exposure kills these cells.
Germline stem cells, with their asymmetrical division and ability to tolerate R2 expression, serve to restore rDNA levels when they get too low. Then the differentiated germ cells serve to weed out mutations—including those introduced during R2 expression in the earlier stem cell stage—by killing off cells with DNA damage. The different strengths of the different types of germ cells creates an effective pipeline to produce the largest number of sperm cells with high rDNA repeat number and low DNA damage.
Eventually, this new understanding of the details of how cells maintain their rDNA could lead to medical therapies. For example, cancer cells are, like germ cells, an essentially immortal cell line, and so must have a way to maintain their rDNA. If researchers could someday find a way to prevent them from doing so, that could be a good treatment strategy. The work also may have implications for research on aging, as rDNA decreases with age in other cell types. In the meantime, Yamashita and Nelson are excited to have solved several long-standing mysteries in their field, including how germ cells can restore rDNA at a population level when each division creates an equal loss and gain of rDNA, and why germline stem cells divide asymmetrically.
“Typically, when you publish a paper, you feel like you’ve fit two puzzle pieces together, but in this case, I feel like we fit a bunch of puzzle pieces together,” Yamashita says. “It’s been immensely satisfying to find answers to multiple questions and see how they all fit together to explain the mechanisms of this process that’s necessary for germline immortality.”